
Journal of Geographical Sciences >
Variation of gross primary productivity dominated by leaf area index in significantly greening area
|
Chen Xin (1996-), PhD, specialized in ecological climatology. E-mail: xin.chen19960607@gmail.com |
Received date: 2022-10-12
Accepted date: 2023-04-11
Online published: 2023-08-29
Supported by
National Natural Science Foundation of China(52078237)
The leaf area index (LAI) shows a significant increasing trend from global to regional scales, which is known as greening. Greening will further enhance photosynthesis, but it is unclear whether the contribution of greening has exceeded the CO2 fertilization effect and become the dominant factor in the gross primary productivity (GPP) variation. We took the Yangtze River Delta (YRD) of China, where cropland and natural vegetation are significantly greening, as an example. Based on the boreal ecosystem productivity simulator (BEPS) and Revised-EC-LUE models, the GPP in the YRD from 2001 to 2020 was simulated, and attribution analysis of the interannual variation in GPP was performed. In addition, the reliability of the GPP simulated by the dynamic global vegetation model (DGVM) in the area was further investigated. The research results showed that GPP in the YRD had three significant characteristics consistent with LAI: (1) GPP showed a significant increasing trend; (2) the multiyear mean and trend of natural vegetation GPP were higher than those of cropland GPP; and (3) cropland GPP showed double-high peak characteristics. The BEPS and Revised-EC-LUE models agreed that the effect of LAI variation (4.29 Tg C yr-1 for BEPS and 2.73 Tg C yr-1 for the Revised-EC-LUE model) determined the interannual variation in GPP, which was much higher than the CO2 fertilization effect (2.29 Tg C yr-1 for BEPS and 0.67 Tg C yr-1 for the Revised-EC-LUE model). The GPP simulated by the 7 DGVMs showed a huge inconsistency with the GPP estimated by remote sensing models. The deviation of LAI simulated by DGVM might be a potential cause for this phenomenon. Our study highlights that in significant greening areas, LAI has dominated GPP variation, both spatially and temporally, and DGVM can correctly simulate GPP only if it accurately simulates LAI variation.
CHEN Xin , CAI Anning , GUO Renjie , LIANG Chuanzhuang , LI Yingying . Variation of gross primary productivity dominated by leaf area index in significantly greening area[J]. Journal of Geographical Sciences, 2023 , 33(8) : 1747 -1764 . DOI: 10.1007/s11442-023-2151-5
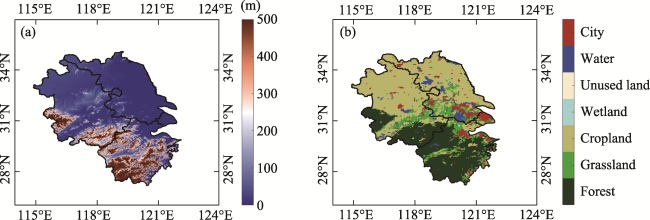
Figure 1 Elevation (a) and land use type (b) from MODIS in 2010 of the Yangtze River Delta |
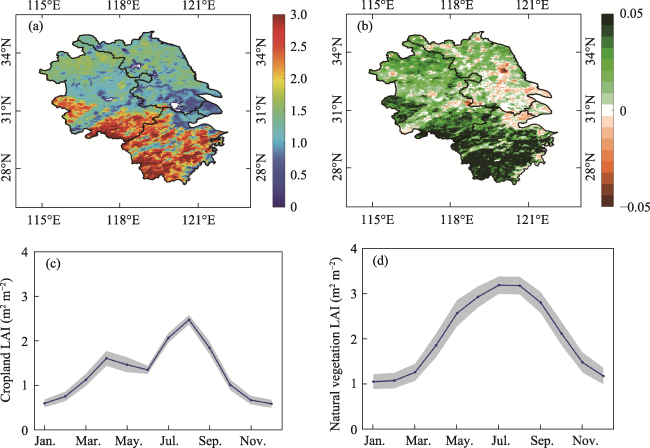
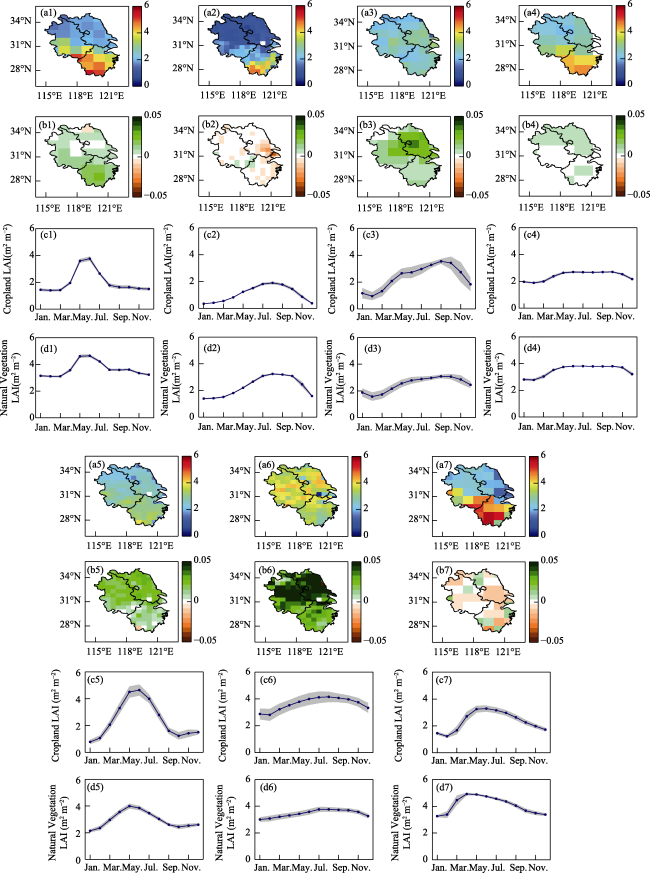
Figure 2 The spatial distribution and seasonal variation in LAI in the Yangtze River Delta from 2001 to 2020. a and b represent the multiyear mean and trend of LAI, c and d represent the LAI seasonal variation in cropland and natural vegetation, the blue line is the mean from 2001 to 2020, and the gray area is the standard deviation. |
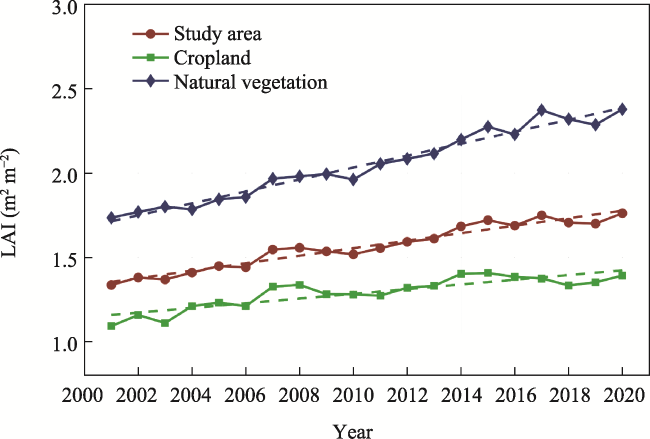
Figure 3 Interannual variation in LAI in the Yangtze River Delta from 2001 to 2020 |
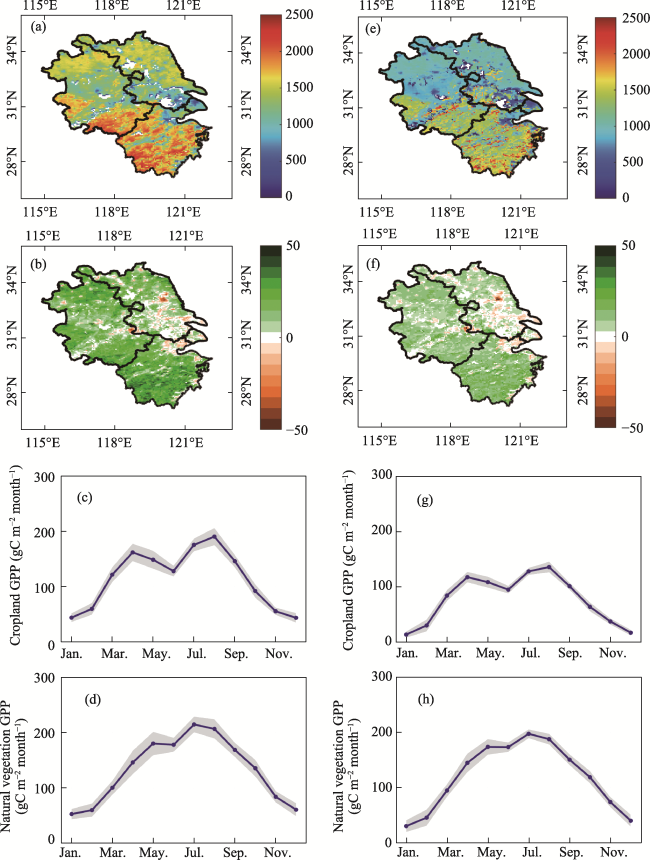
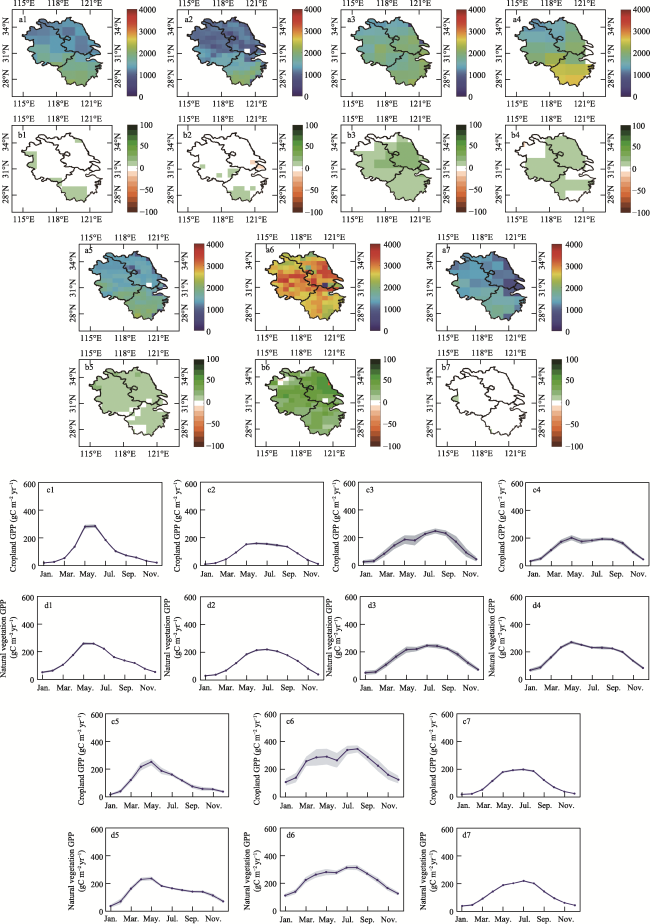
Figure 4 Spatial distribution and seasonal variation in GPP simulated by BEPS and the Revised-EC-LUE model in the Yangtze River Delta. a-d represent the multiyear mean, trend, and seasonal variation in GPP of cropland and natural vegetation simulated by BEPS from 2001 to 2020, e-h represent the multiyear mean, trend, and seasonal variation in GPP of cropland and natural vegetation simulated by the Revised-EC-LUE model from 2001 to 2020, the blue line is the mean from 2001 to 2020, and the gray area is the standard deviation. |
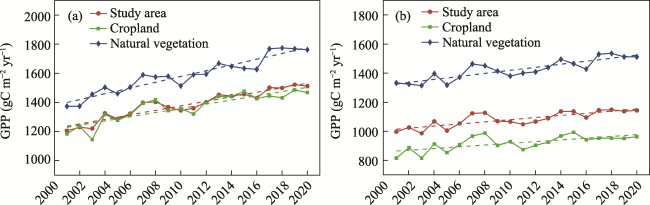
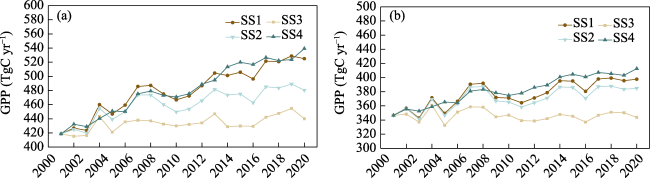
Figure 5 Interannual variation in GPP simulated by BEPS (a) and the Revised EC-LUE model (b) in the Yangtze River Delta from 2001 to 2020 |
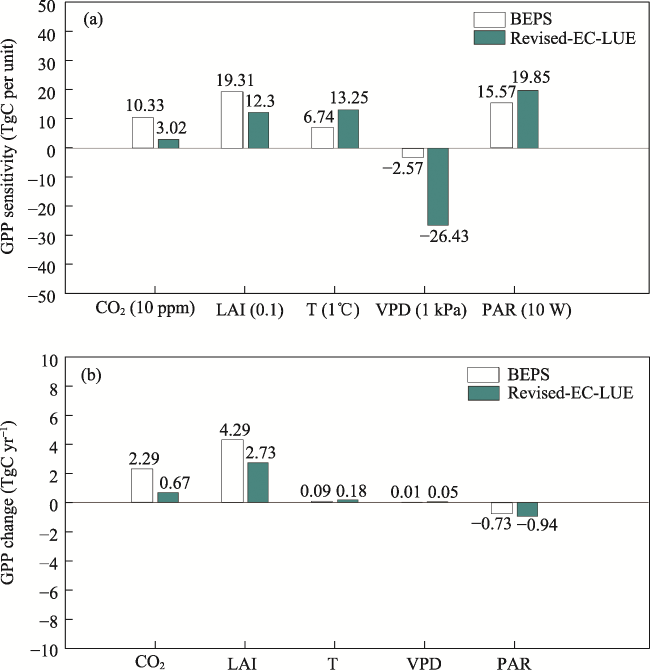
Figure 6 Sensitivity analysis of the drivers of GPP variation in the Yangtze River Delta from 2001 to 2020 (a) and the actual impact of drivers on GPP variation in the Yangtze River Delta (b) |
Figure 7 The spatial distribution and seasonal variation in GPP simulated by 7 DGVMs in the Yangtze River Delta. a and b represent the multiyear mean and trend of GPP; c and d represent the seasonal variation in GPP of cropland and natural vegetation, the blue line is the mean from 2001 to 2019, and the gray area is the standard deviation. 1-7 are GLM5.0, ISAM, ISBA-CTRIP, JULES-ES, LPJ-LUESS, ORCHIDEEv3, and SDGVM, respectively. |
Table 1 Multiyear mean and trend of GPP in the Yangtze River Delta simulated by 7 DGVMs (gC m-2 yr-1 for average and gC m-2 yr-2 for trend) |
| Study area | Cropland | Natural vegetation | ||||
|---|---|---|---|---|---|---|
| Average | Trend | Average | Trend | Average | Trend | |
| CLM5.0 | 1.48×103 | 4.71* | 1.3×103 | 4.32* | 1.7×103 | 5.25* |
| ISAM | 1.25×103 | 1.85* | 1.09×103 | 0.89 | 1.5×103 | 3.6* |
| ISBA-CTRIP | 1.79×103 | 13.37* | 1.69×103 | 13.13 | 1.88×103 | 12.74* |
| JULES-ES | 1.87×103 | 6.87* | 1.61×103 | 6.15 | 2.19×103 | 7.68* |
| LPJ-GUESS | 1.51×103 | 11.05* | 1.36×103 | 13.61* | 1.73×103 | 8.49* |
| ORCHIDEEv3 | 2.76×103 | 33.78* | 2.85×103 | 37.18* | 2.74×103 | 30.43* |
| SDGVM | 1.32×103 | 1.62 | 1.23×103 | 1.04* | 1.47×103 | 2.73 |
Note: * indicates p < 0.05. |
Table 2 The trend of GPP simulated by the 7 DGVMs under different simulation scenarios and the response of GPP to different variables in the Yangtze River Delta (Tg C yr-1) |
| S0 | S1 | S2 | S3 | CO2 | Climate | LUCC | |
|---|---|---|---|---|---|---|---|
| CLM5.0 | 0 | 1.91* | 1.98* | 1.64* | 1.91 | 0.07 | -0.35 |
| ISAM | -0.19 | 1.57* | 1.53* | 0.64* | 1.77* | -0.05 | -0.88* |
| ISBA-CTRIP | -0.37 | 1.59 | 3.73 | 4.65* | 1.97* | 2.14 | 0.92* |
| JULES-ES | -0.38 | 1.72 | 1.84 | 2.39* | 2.1* | 0.12 | 0.54 |
| LPJ-GUESS | -0.07 | 1.94* | 2.03* | 3.84* | 2.01* | 0.09 | 1.81* |
| ORCHIDEEv3 | -0.19 | 3* | 4.68* | 11.75* | 3.19* | 1.68* | 7.06* |
| SDGVM | -0.92* | 0.23 | 2.27* | 0.56 | 1.15* | 2.04* | -1.7* |
Note: * indicates p < 0.05. |
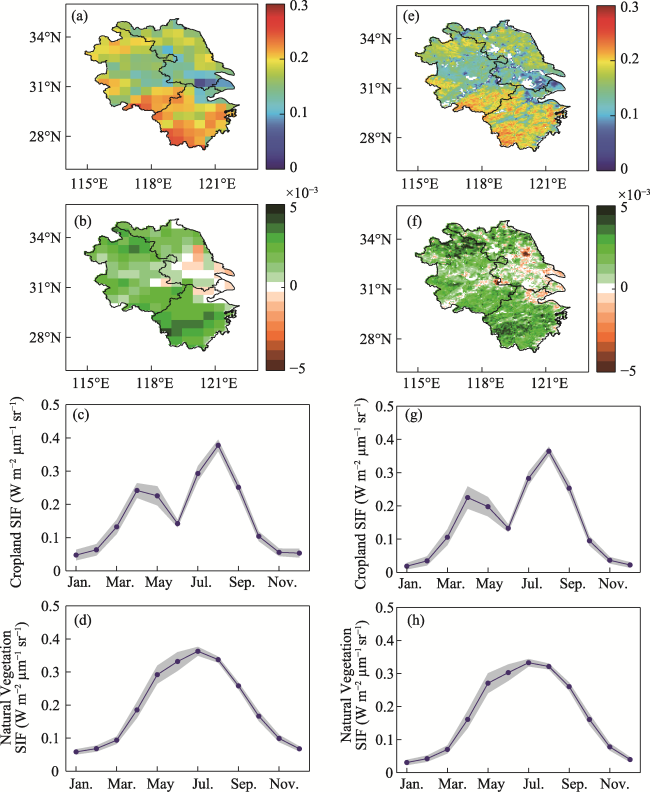
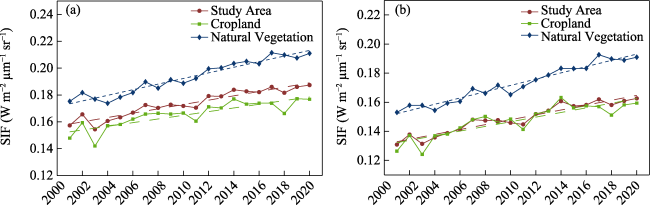
Figure S1 Spatial distribution and seasonal variation in CSIF and GOSIF. a-d represent the multiyear mean, trend, and seasonal variation in cropland and natural vegetation of CSIF from 2001 to 2020, e-h represent the multiyear mean, trend, seasonal variation in cropland and natural vegetation of GOSIF from 2001 to 2020, the blue line is the mean from 2001 to 2020, and the gray area is the standard deviation. |
Figure S2 Interannual variation in CSIF (a) and GOSIF (b) from 2001 to 2020 |
Figure S3 Variation in GPP in the YRD from 2001 to 2020 simulated by BEPS (a) and the Revised-EC-LUE model (b) under different simulation scenarios |
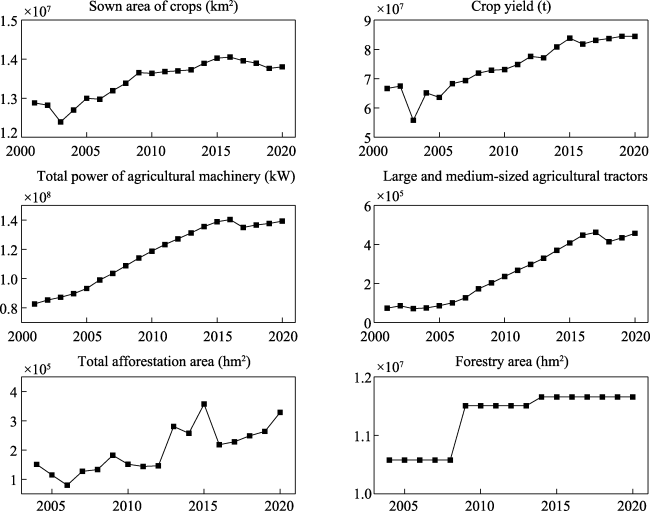
Figure S4 Variations in some agriculture and forestry in the Yangtze River Delta. The temporal range of agriculture is 2001-2020, and the time range of forestry is 2004-2020 |
Figure S5 The spatial distribution and seasonal variation in LAI simulated by 7 DGVMs in the Yangtze River Delta. a and b represent the multiyear mean and trend of LAI, c and d represent the seasonal variation in LAI of cropland and natural vegetation, the blue line is the mean from 2001 to 2019, and the gray area is the standard deviation. 1-7 are GLM5.0, ISAM, ISBA-CTRIP, JULES-ES, LPJ-LUESS, ORCHIDEEv3, and SDGVM, respectively. |
Table S1 Multiyear mean and trend of LAI simulated by 7 DGVMs in the Yangtze River Delta (m2 m-2 for average and m2 m-2 yr-1 for trend) |
| Study area | Cropland | Natural vegetation | ||||
|---|---|---|---|---|---|---|
| Average | Trend | Average | Trend | Average | Trend | |
| CLM5.0 | 2.76 | 7.92×10-3* | 2.02 | 4.4×10-3 | 3.65 | 12.19×10-3* |
| ISAM | 1.62 | -2.6×10-3 | 1.09 | -2.7×10-3 | 2.3 | -1.37×10-3 |
| ISBA-CTRIP | 2.46 | 13.45×10-3 | 2.39 | 14.47×10-3 | 2.5 | 11.15×10-3 |
| JULES-ES | 2.89 | 3.2×10-3* | 2.42 | 3.76×10-3* | 3.48 | 2.6×10-3* |
| LPJ-GUESS | 2.63 | 16.53×10-3* | 2.41 | 21.39×10-3* | 2.96 | 11.22×10-3* |
| ORCHIDEEv3 | 3.49 | 36.34×10-3* | 3.63 | 43.04×10-3* | 3.45 | 29.52×10-3* |
| SDGVM | 3.1 | -3.12×10-3 | 2.36 | -2.85×10-3 | 4.1 | -2.88×10-3 |
Note: * indicates p < 0.05. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}