
Journal of Geographical Sciences >
Understanding the spatial heterogeneity of grazing pressure in the Three-River-Source Region on the Tibetan Plateau
|
Gu Changjun (1992-), specialized in grassland change, climate change, and grazing management. E-mail: gucj0308@gmail.com |
Received date: 2022-07-05
Accepted date: 2023-02-23
Online published: 2023-08-29
Supported by
The Second Tibetan Plateau Scientific Expedition and Research(2019QZKK0603)
Strategic Priority Research Program of the Chinese Academy of Sciences(XDA20040201)
National Natural Science Foundation of China(41671104)
Elucidating the distribution of the grazing pressure requires an understanding of the grazing activities. In this study, we analyzed the grazing behavior of yaks in Three-River- Source Region (TRSR) and identified the main factors influencing the distribution of grazing intensity (GI) using trajectory data and remote sensing datasets. Our results revealed that a semi-resident transhumance strategy is employed in this region. The average grazing time (GT) of four GPS collars over the year was 11.84 h/day (N6), 11.01 h/day (N11), 9.25 h/day (N18), and 11.61 h/day (N24). GT was generally higher in warm seasons (summer and autumn) than in cold seasons (spring and winter). The average daily moving speed was found to be closely related to the pasture size of different herders and the seasons. Geodetector analysis identified the distance to camp (DOC) as the most important single factor influencing the distribution of GI, explaining up to 52% of the GI variations. However, relying solely on this factor may not accurately depict the actual GI distribution. When pairwise factors interacted, the explanatory power of the model increased, ranging from 34.55% to 63.26%. Our study highlights the importance of considering multiple factors when predicting grazing intensity, as grazing activities tend to cluster near settlements, but other factors may also be influential.
Key words: grazing intensity; GPS-tracking; spatial heterogeneity; yak; Tibetan Plateau
GU Changjun , LIU Linshan , ZHANG Yili , WEI Bo , CUI Bohao , GONG Dianqing . Understanding the spatial heterogeneity of grazing pressure in the Three-River-Source Region on the Tibetan Plateau[J]. Journal of Geographical Sciences, 2023 , 33(8) : 1660 -1680 . DOI: 10.1007/s11442-023-2147-1
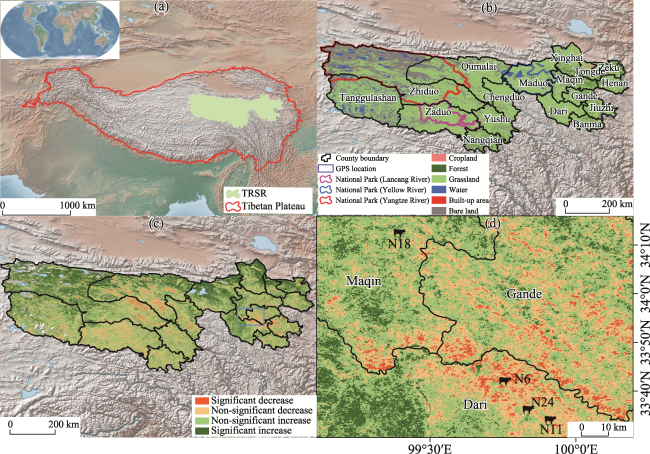
Figure 1 Three-River-Source Region (TRSR) is located in central Tibetan Plateau (TP) (Zhang et al., 2021) and has a first national park in China and also the biggest natural reserve in China (a). As a typical alpine ecosystem, TRSR is characterized by alpine grassland and alpine meadow (b). The Chinese government has implemented ecological conservation and restoration project in this region. Since the beginning of the 21st century, the greenness of vegetation has maintained positive improvement generally but decreased locally (c). Four GPS tracking devices were set in Maqin and Dari county (d). |
Figure 2 Diagram of GPS collar used in this study (a): ① A Nylon collar; ② A screwdriver; ③ Waterproof box; ④⑤ USB cable; ⑥ Instructions; ⑦ GPS tracker; ⑧ Packing box; Selected yak with GPS collar in Dari county, located in central TRSR (b) |
Table 1 List of GPS location records of grazing yaks during the past year (August 2020 to September 2021) |
| Month | N6 | N11 | N18 | N24 |
|---|---|---|---|---|
| 1 | 16724 | 14802 | 6535 | 17694 |
| 2 | 20826 | 16521 | 15066 | 16944 |
| 3 | 16097 | 16814 | 16008 | 17919 |
| 4 | 14956 | 13081 | 16736 | 14977 |
| 5 | 24786 | 15124 | 24575 | 19966 |
| 6 | 19404 | 14783 | 20521 | 21365 |
| 7 | 18598 | 17251 | 12824 | 18627 |
| 8 | 13130 | 22483 | 17355 | 21545 |
| 9 | 10021 | 20469 | 13910 | 17042 |
| 10 | 7909 | 8894 | 17366 | 18087 |
| 11 | 2945 | 11916 | 17280 | 15958 |
| 12 | 17458 | 14679 | 18456 | 16100 |
| Total | 182854 | 186817 | 196632 | 216224 |
Table 2 A detail time table of grazing activities across the four seasons |
| Spring | Summer | Autumn | Winter | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Avg | Min | Max | Avg | Min | Max | Avg | Min | Max | Avg | Min | Max | |
| Start | 10:16 | 6:38 | 17:12 | 8:08 | 5:58 | 14:27 | 9:40 | 7:43 | 12:26 | 11:26 | 10:29 | 13:57 |
| End | 19:52 | 11:21 | 22:39 | 20:48 | 19:18 | 23:05 | 19:55 | 18:02 | 21:19 | 18:33 | 11:00 | 20:13 |
| GT | 9:36 | 12:40 | 10:15 | 7:07 | ||||||||
Table 3 Potential environmental factors influencing the distribution of grazing intensity |
| Data source | Resolution | Year | |
|---|---|---|---|
| Elevation | NASA SRTM Digital Elevation 30 m, https://cmr.earthdata.nasa.gov/search | 30 m×30 m | 2000 |
| Slope | Obtained from DEM | 30 m×30 m | 2000 |
| Aspects | Obtained from DEM | 30 m×30 m | 2000 |
| NDVI | Landsat 8 Collection 1 Tier 1 32-Day NDVI Composite, https://earthengine.google.com/ | 30 m×30 m | 2016-2021 (Monthly average) |
| Distance to camp (DOC) | Digitization of google earth images | —— | |
| Distance to road (DOD) | Digitization of google earth images | —— | |
| Distance to river (DOR) | Digitization of google earth images | —— | |
| Month | —— | —— |
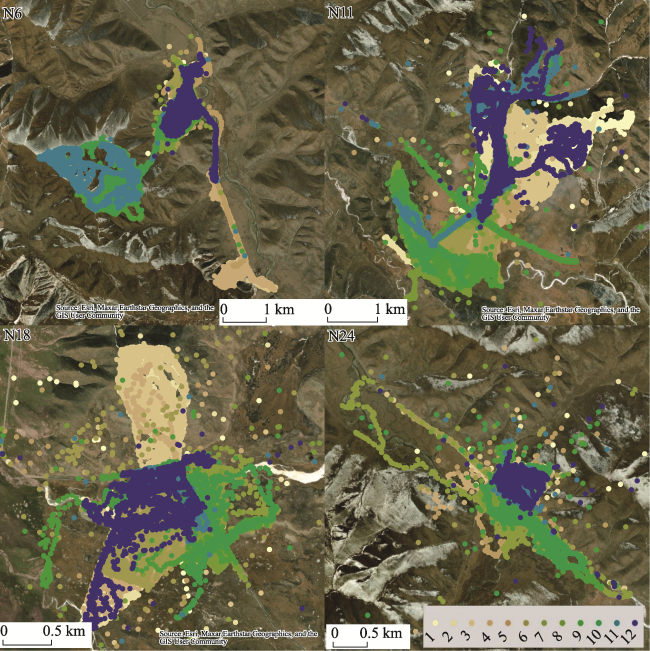
Figure 3 Spatial patterns of GPS tracks during the whole year (August 2020 to September 2021) in four locations |
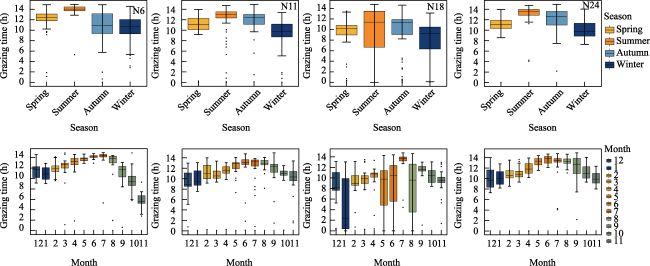
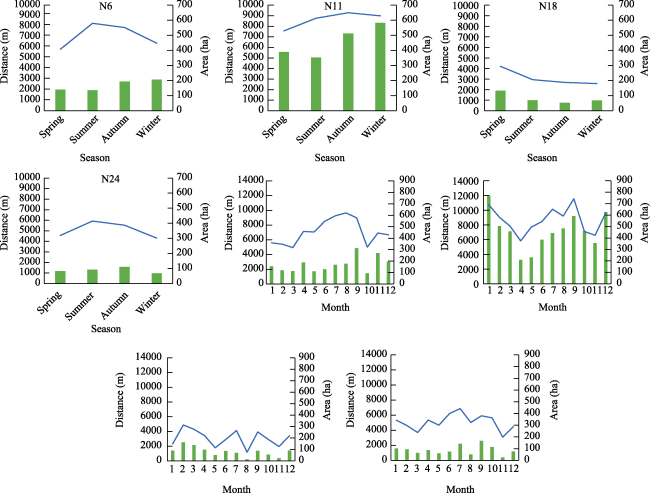
Figure 4 Grazing time (GT) in different seasons and months of four GPS collars during the past whole year (August 2020 to September 2021) |
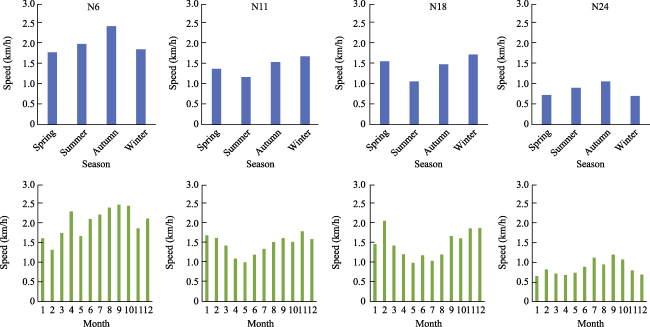
Figure 5 Moving speed of yaks in four GPS collars in different seasons and months |
Figure 6 Average daily moving distance and home range of yaks in different seasons and months |
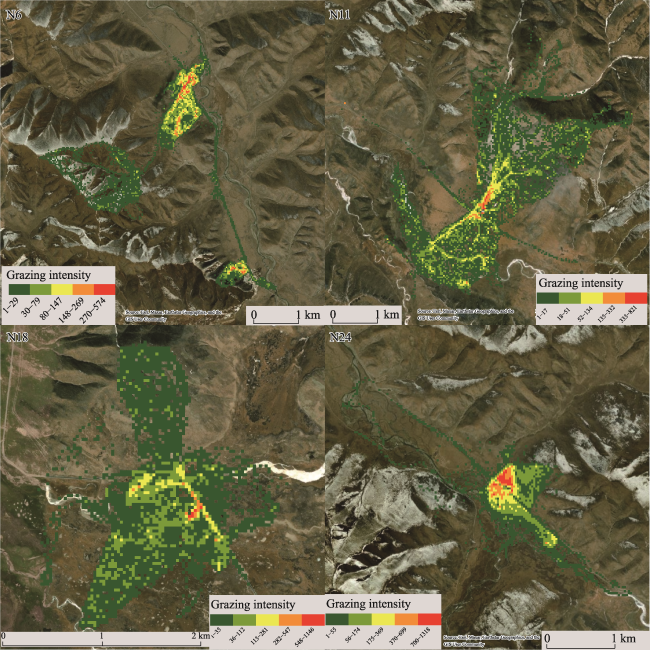
Figure 7 Distribution of grazing intensity of four GPS collars |
Table 4 The q-statistics value of seven main influencing factors of grazing intensity |
| q-statistics | Elevation | Slope | Aspect | NDVI | Distance to camp (DOC) | Distance to road (DOD) | Distance to river (DOR) | Month |
|---|---|---|---|---|---|---|---|---|
| N6 | 0.16 | 0.02 | 0.02 | 0.09 | 0.12 | 0.15 | 0.01 | 0.34 |
| N11 | 0.10 | 0.04 | 0.01 | 0.03 | 0.19 | 0.07 | 0.04 | 0.05 |
| N18 | 0.02 | 0.01 | 0 | 0.02 | 0.20 | 0.08 | 0.03 | 0.05 |
| N24 | 0.03 | 0.04 | 0.02 | 0.09 | 0.52 | 0.13 | 0.04 | 0.25 |
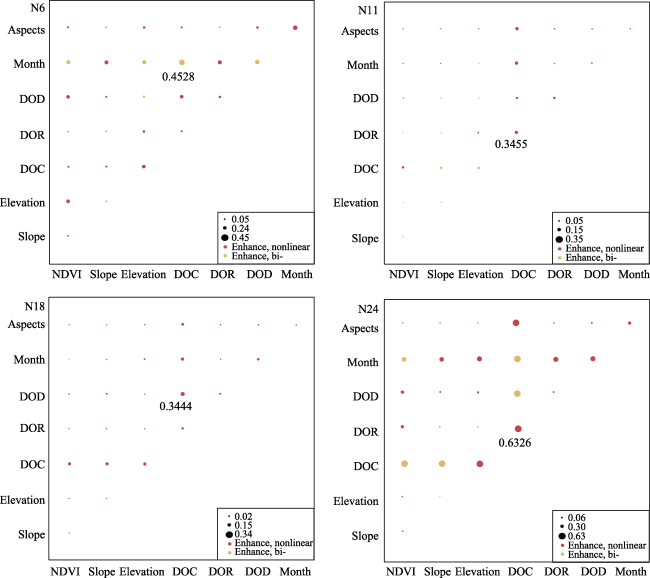
Figure 8 Interactions (measured by q value) between pairs of factors on grazing intensity distribution. DOC, DOR, and DOD represent distance to camp, distance to river, and distance to road, respectively. |
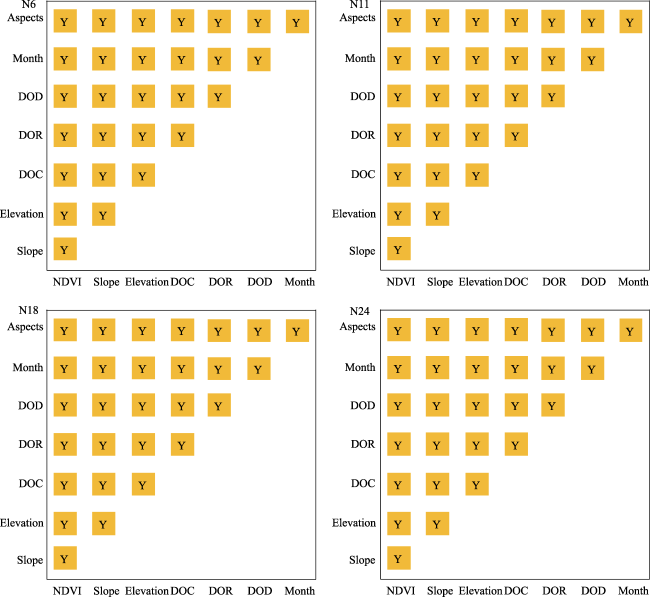
Figure 9 Statistical significance of the pairwise factors influences on grazing intensity distribution (95% confidence level). DOC, DOR, and DOD represent distance to camp, distance to river, and distance to road, respectively. Y indicates that there are significant differences existed in the effects of the pairwise factors on grazing intensity distribution. |
Table 5 Corresponding range of eight factors to maximum grazing intensity |
| Elevation (m) | Slope (°) | Aspect | NDVI | DOC (m) | DOR (m) | DOD (m) | Month | |
|---|---|---|---|---|---|---|---|---|
| N6 | 4000-4030 | 3.68-7.81 | 20-73.7 | 0.16-0.24 | 12.3-270 | 653-816 | 42.7-138 | 1 |
| N11 | 4080-4130 | 1.85-2.9 | 178-237 | 0.41-0.55 | 40.8-87.2 | 0.04-44.9 | 0.07-321 | 6 |
| N18 | 4258-4261 | 3.57-4.48 | 182-243 | 0.08-0.17 | 28.9-65.9 | 252-336 | 0.07-3.58 | 1 |
| N24 | 4033-4041 | 4.98-9.25 | 211-236 | 0.16-0.26 | 49.8-112 | 112-194 | 40.4-107 | 1 |
Note: DOC, DOR, and DOD represent the distance to camp, the river, and the road, respectively. |

Figure 10 Routine corridors of grazing activities |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}