
Journal of Geographical Sciences >
Late Pleistocene vegetation succession, climate change and hominin adaptation in Sandinggai site, central South China
|
Lu Lili (1993-), PhD, specialized in environmental archaeology. E-mail: lulili@ivpp.ac.cn |
Received date: 2024-09-03
Accepted date: 2025-02-12
Online published: 2025-09-04
Supported by
National Natural Science Foundation of China(42471185)
National Natural Science Foundation of China(T2192952)
National Key Research and Development Program of China(2022YFF0801502)
The paleoenvironmental changes and adaptation strategies of hominins during the Late Pleistocene are crucial for understanding the evolution, dispersal, and behavioral shifts of early modern humans. Despite South China’s significance as a nexus for hominin dispersal and handaxe technology diffusion, quantitative reconstructions of paleoenvironments linked to archaeological records remain scarce. The Sandinggai site (96.6-13.3 ka BP) in central South China, with its well-preserved stratigraphy and abundant lithic artefacts, is notable for providing valuable insights. In this study, quantitative reconstruction of the vegetation succession and climate change sequences at the site was conducted using palynological and isotopic data. The results indicated a shift from a warm-temperate evergreen and deciduous broadleaf mixed forest to a temperate deciduous broadleaf forest, with the climate transitioning from warm and humid to cooler and drier conditions. During the early phase, an increase in lithic production suggested favorable conditions for hominin survival. In the later phase, decreased lithic production and the replacement of large handaxe tools by smaller flake tools, indicated that hominins adapted to the cooler, drier climate and more open landscapes through lithic miniaturization. These findings highlight the environment-driven adaptation of lithic technology and hominin behavior, thereby shedding light on human survival adaptation strategies.
LU Lili , ZHAO Keliang , LI Yiyuan , LI Hao , LIU Junchi , BAI Guangyi , XIAO Peiyuan , YANG Qingjiang , LI Xiaoqiang . Late Pleistocene vegetation succession, climate change and hominin adaptation in Sandinggai site, central South China[J]. Journal of Geographical Sciences, 2025 , 35(8) : 1642 -1666 . DOI: 10.1007/s11442-025-2387-3
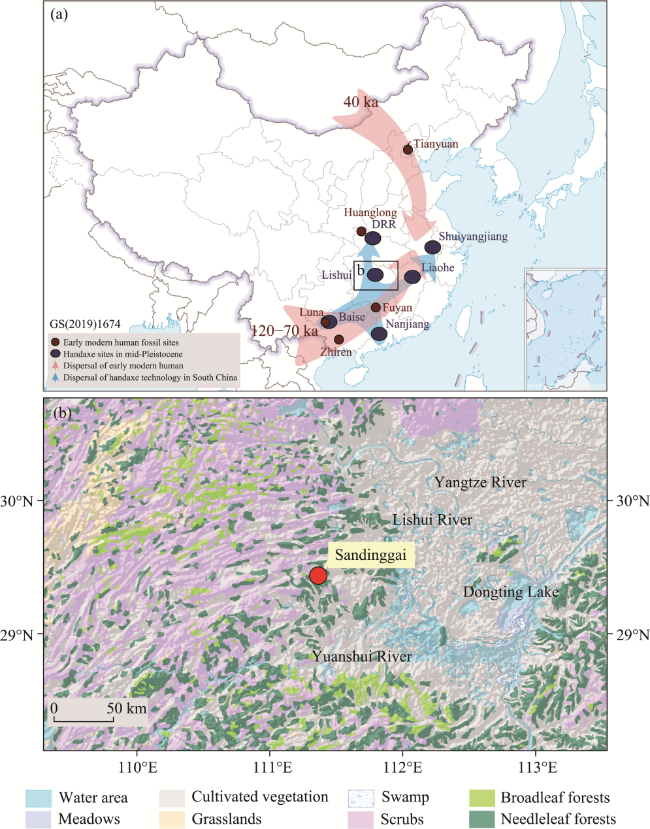
Figure 1 Distribution of early modern humans and handaxe technology in southern China and the geographical location of the Sandinggai site (a. Red dots indicate representative fossil sites of early modern humans in China during the Late Pleistocene (Shang et al., 2007; Liu et al., 2010; Liu et al., 2015; Bae, 2017), and red arrows represent their possible diffusion routes (Bae et al., 2017). Blue ovals indicate representative sites of handaxe technology from the Middle to Late Pleistocene in southern China (Li and Xu, 1991; Hou et al., 2000; Dong et al., 2019; Li et al., 2022; Dai, 2023; Li and Song, 2023), with blue arrows showing their potential dispersal routes (Li et al., 2024); b. Geographic location of the Sandinggai site, displaying the surrounding topography and vegetation (Editorial Committee of China Vegetation Map CAS, 2001).) |
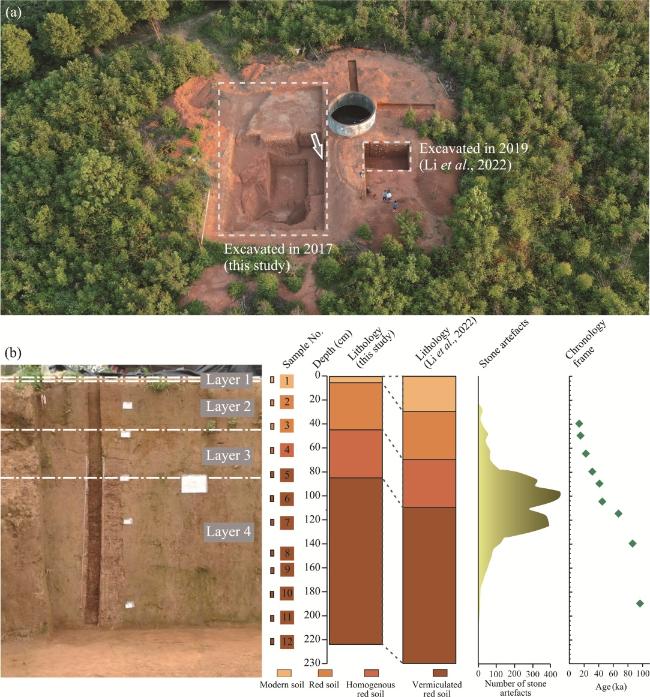
Figure 2 Basic information of sampling at the Sandinggai site (a. Location of excavated trenches, modified from Li et al. (2022). The arrow indicates the sampling section of this study. b. Stratigraphy, lithology, pollen samples, lithic artefact distribution, and chronological sequence (Li et al., 2022).) |
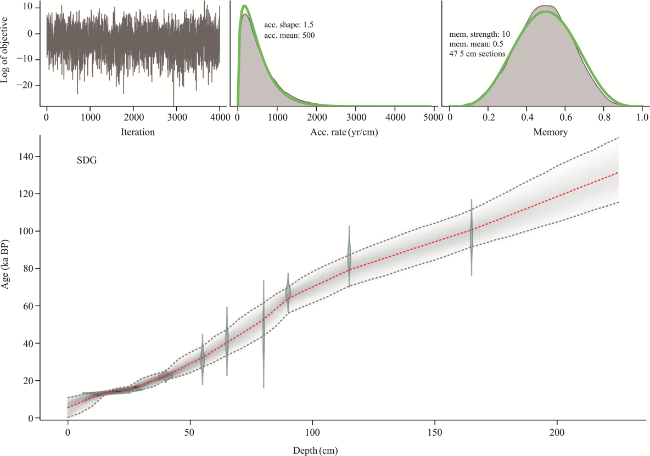
Figure 3 Age framework of pollen samples calculated using the Bacon age-depth model |
Table 1 Depth, age, pollen counts and pollen concentration data for each sample from Sandinggai section |
| Sample No. | Depth (cm) | Age (ka) | Sum (grains) | Concentration (grains/g) | |
|---|---|---|---|---|---|
| Layer 1 (0-5 cm) | 1 | 5 | 9.355 | 436 | 4034 |
| Layer 2 (5-45 cm) | 2 | 25 | 15.313 | 341 | 32 |
| 3 | 45 | 25.808 | 427 | 31 | |
| Layer 3 (45-85 cm) | 4 | 65 | 40.5 | 144 | 7 |
| 5 | 85 | 58.219 | 80 | 3 | |
| Layer 4 (85-225 cm) | 6 | 105 | 73.098 | 134 | 5 |
| 7 | 125 | 83.486 | 189 | 4 | |
| 8 | 150 | 94.172 | 107 | 3 | |
| 9 | 165 | 100.53 | 428 | 16 | |
| 10 | 185 | 110.489 | 70 | 2 | |
| 11 | 205 | 120.67 | 37 | 1 | |
| 12 | 225 | 130.921 | 35 | 1 |
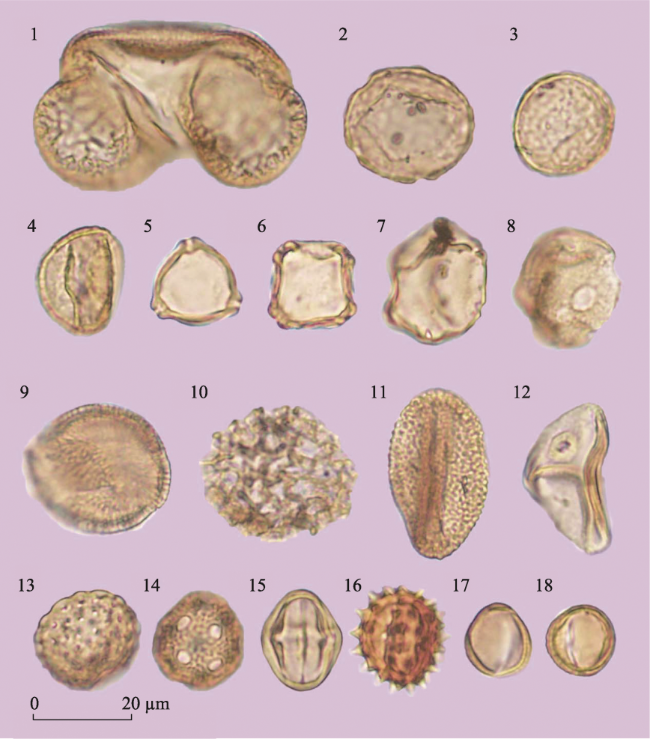
Figure 4 Major palynomorphs from the Sandinggai section (1. Pinus; 2-3. Ulmus; 4. Quercus; 5. Betula; 6. Alnus; 7. Juglans; 8. Liquidambar; 9. Hamamelidaceae; 10. Polygonum; 11. Fagopyrum; 12. Poaceae; 13. Chenopodiaceae; 14. Caryophyllaceae; 15. Rosaceae; 16. Asteraceae (excl. Artemisia); 17-18. Artemisia) |
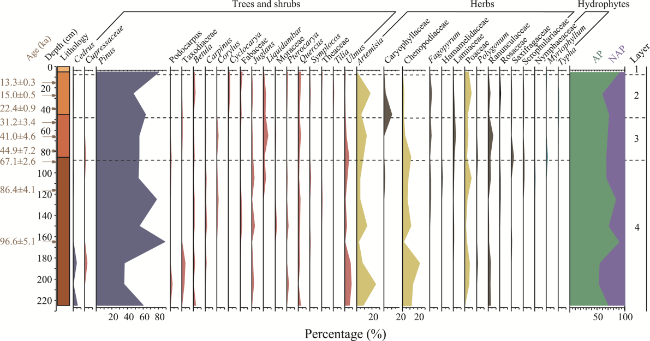
Figure 5 Diagram showing changes in the relative abundances of the major palynomorphs recovered from the Sandinggai section |
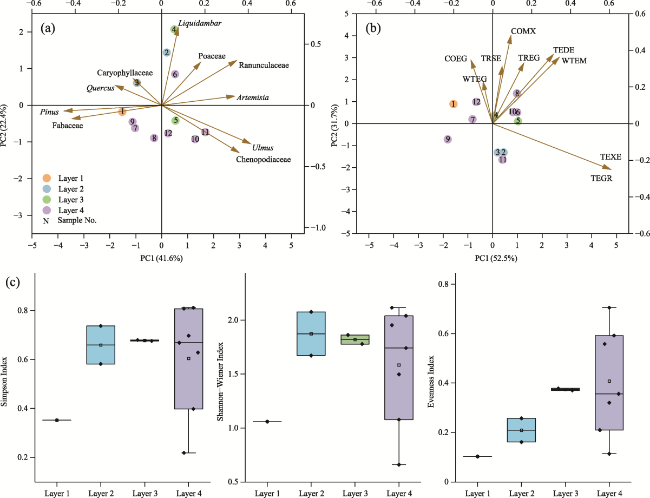
Figure 6 Palaeoenvironmental analyses of palynomorphs, biomes and plant diversity data from the Sandinggai section (a. Principal component analysis of 12 pollen samples and 10 palynomorphs with relative abundance greater than 1%; b. Principal component analysis of 12 pollen samples and 9 major biomes; c. Box plot showing the distribution of plant diversity indices (Simpson Index, Shannon-Wiener Index and Evenness Index) across four layers) |
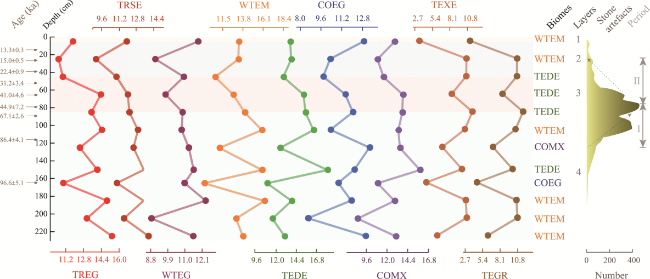
Figure 7 Biome score sequences and vegetation reconstruction of the Sandinggai site (TREG: tropical evergreen broadleaf forest; TRSE: tropical semi-evergreen broadleaf forest; WTEG: warm-temperate evergreen broadleaf forest; WTEM: warm-temperate evergreen broadleaf and mixed forest; TEDE: temperate deciduous broadleaf forest; COMX: cool mixed forest; COEG: cool evergreen needleleaf forest; TEXE: temperate xerophytic shrubland; TEGR: temperate grassland. Period I: a phase of increasing lithics; Period II: a phase of decreasing lithics) |
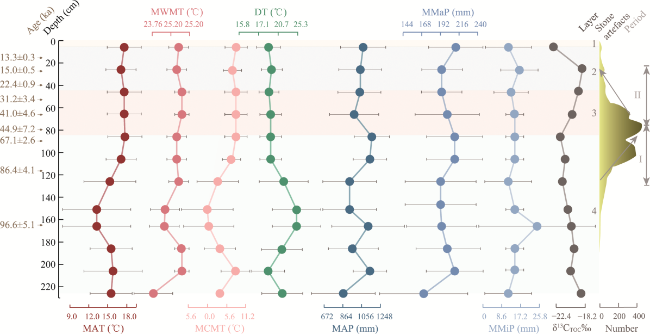
Figure 8 Changes in seven climatic parameters and total organic carbon isotopes at the Sandinggai site (MAT: the mean annual temperature; MWMT: the mean warmest monthly temperature; MCMT: the mean coldest monthly temperature; DT: the temperature difference between the coldest and warmest months; MAP: the mean annual precipitation; MMaP: the mean maximum monthly precipitation; MMiP: the mean minimum monthly precipitation. Period I: a phase of increasing lithics; Period II: a phase of decreasing lithics) |
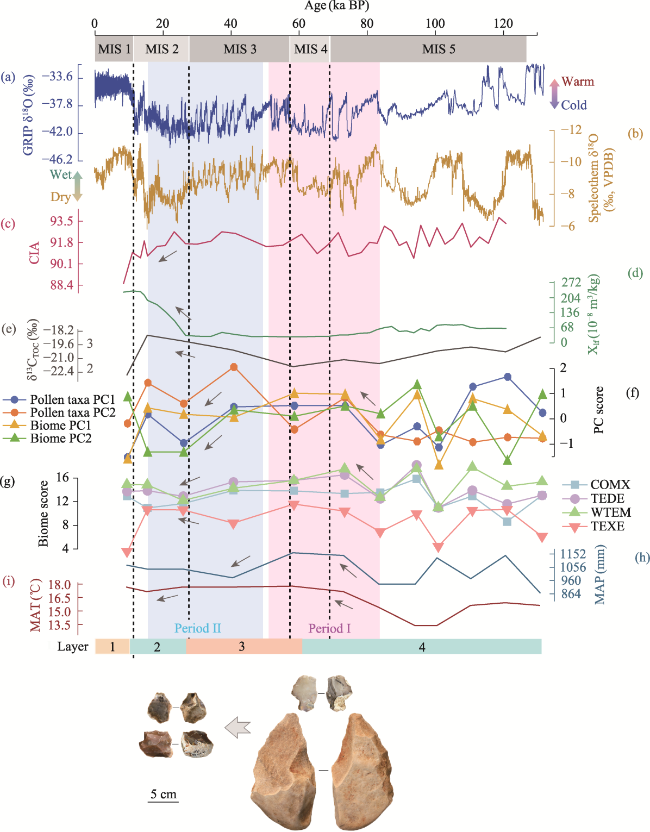
Figure 9 Multidisciplinary indicators for comparing palaeoenvironmental changes at the Sandinggai site (a. Late Pleistocene climatic change recorded in the Greenland Ice Core Project (GRIP, Johnsen et al., 1997); b. Late Pleistocene climatic change documented in Chinese speleothems at the Sanbao, Hulu and Dongge caves (Cheng et al., 2018); c. CIA (Li et al., 2022); d. Magnetic susceptibility (Li et al., 2022); e. Total organic carbon isotope (this study); f. PC value (this study); g. Main biome scores (this study); h. MAP (this study); i. MAT (this study)) |
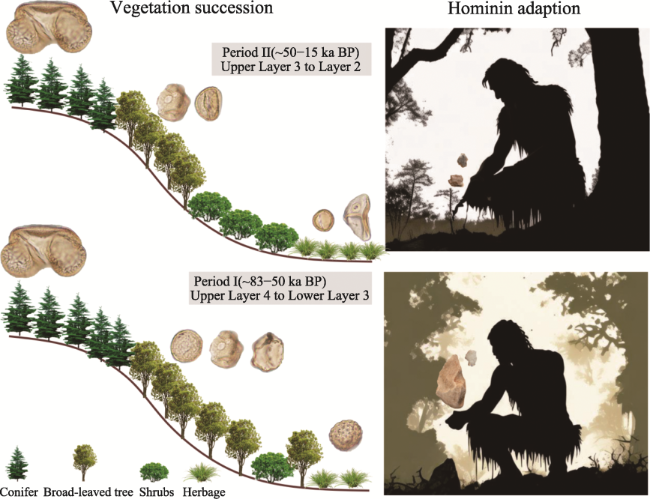
Figure 10 Vegetation succession and hominin adaptations at the Sandinggai site during the Late Pleistocene. The left column schematically represents vegetation succession, while the right column shows the adaptation of stone artefacts used by hominins in response to increasingly open vegetation. The background images in the right column are sourced from AI Creations (https://image.baidu.com/). |
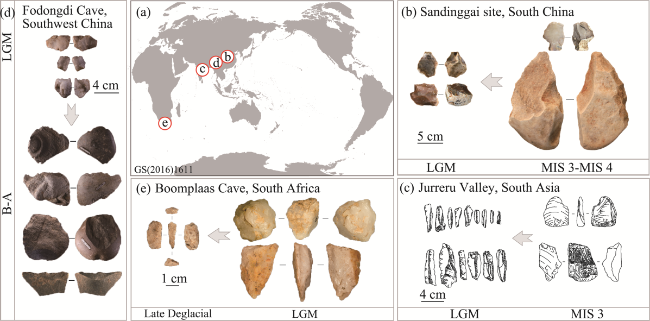
Figure 11 Snapshots of lithic miniaturisation at representative sites globally around the LGM (a. Site distribution; b. Sandinggai site (Li et al., 2022); c. Jurreru Valley, South Asia (Petraglia et al., 2009); d. Fodongdi Cave (Huan et al., 2024); e. Boomplaas Cave, South Africa (Pargeter and Faith, 2020)) |
Table S1 Plant function types (PFTs) and the pollen taxa assigned to them (Prentice et al., 1996; Tarasov et al., 1998; Ni et al., 2010) |
| Codes | Plant functional type | Main pollen taxa included | ||
|---|---|---|---|---|
| wt.e.n.t | Warm-temperate evergreen needle-leaved tree | Cedrus, Pinus, Podocarpus, Taxodiaceae, Tsuga | ||
| eu.e.n.t | Eurythermic evergreen needle-leaved tree | Cupressaceae, Pinus | ||
| wt.d.n.t | Warm-temperate deciduous needle-leaved tree | Taxodiaceae | ||
| wt.e.sb.t | Warm-temperate evergreen sclerophyll broad-leaved tree | Euphorbiaceae, Fabaceae, Oleaceae, Quercus | ||
| wt.e.mb.t | Warm-temperate evergreen malacophyll broadleaf tree | Apocynaceae, Araliaceae, Fabaceae, Hamamelidaceae, Moraceae, Oleaceae, Quercus, Scrophulariaceae, Symplocos, Theaceae | ||
| tr.e.sb.t | Tropical evergreen sclerophyll broadleaf tree | Euphorbiaceae, Fabaceae, Oleaceae | ||
| tr.e.mb.t | Tropical evergreen malacophyll broadleaf tree | Apocynaceae, Araliaceae, Euphorbiaceae, Fabaceae, Moraceae, Oleaceae, Scrophulariaceae, Tilia, Ulmus | ||
| te-fa.cd.mb.t | Temperate (spring-frost avoiding) cold-deciduous malacophyll broadleaf tree | Carpinus, Euphorbiaceae, Fabaceae, Lamiaceae, Oleaceae, Quercus, Rosaceae, Scrophulariaceae, Ulmus | ||
| te-ft.cd.mb.t | Temperate (spring-frost tolerant) cold-deciduous malacophyll broadleaf tree | Acer, Alnus, Betula, Corylus, Fabaceae, Quercus, Rosaceae, Tilia, Ulmus | ||
| te-fi.cd.mb.t | Temperate (spring-frost intolerant) cold-deciduous malacophyll broadleaf tree | Acer, Betula, Caprifoliaceae, Carpinus, Carya, Fabaceae, Hamamelidaceae, Juglans, Liquidambar, Moraceae, Pterocarya, Quercus, Rosaceae, Tilia | ||
| wt.cd.mb.t | Warm-temperate cold-deciduous malacophyll broadleaf tree | |||
| tr-m.dd.mb.t | Tropical mesic drought-deciduous malacophyll broadleaf tree | |||
| tr-x.dd.mb.t | Tropical xeric drought-deciduous malacophyll broadleaf tree | Euphorbiaceae, Fabaceae, Tilia | ||
| sl.t | Small-leaved tree | Alnus, Caprifoliaceae, Carya, Cyclocarya, Euphorbiaceae, Fabaceae, Hamamelidaceae, Juglans, Liquidambar, Moraceae, Pterocarya, Rosaceae, Ulmus | ||
| dt.sl.lhs | Drought-tolerant small-leaved low or high shrub | Euphorbiaceae, Fabaceae, Oleaceae, Tilia, Ulmus | ||
| wt.e.sb.lhs | Warm-temperate evergreen sclerophyll broadleaf low or high shrub | Euphorbiaceae, Fabaceae, Hamamelidaceae, Oleaceae, Quercus, Theaceae | ||
| wt.e.mb.lhs | Warm-temperate evergreen malacophyll broadleaf low or high shrub | Apocynaceae, Araliaceae, Caprifoliaceae, Chloranthus, Euphorbiaceae, Fabaceae, Moraceae, Oleaceae, Quercus, Scrophulariaceae, Symplocos, Viburnum | ||
| tr.e.mb.lhs | Tropical evergreen malacophyll broadleaf low or high shrub | Apocynaceae, Araliaceae, Corylus, Fabaceae, Moraceae, Oleaceae, Scrophulariaceae | ||
| te.cd.mb.lhs | Temperate cold-deciduous malacophyll broadleaf low or high shrub | Alnus, Betula, Caprifoliaceae, Carpinus, Euphorbiaceae, Hamamelidaceae, Lamiaceae, Moraceae, Oleaceae, Quercus, Rosaceae, Scrophulariaceae, Tilia, Ulmus, Viburnum | ||
| wt.cd.mb.lhs | Warm-temperate cold-deciduous malacophyll broadleaf low or high shrub | Caprifoliaceae, Euphorbiaceae, Fabaceae, Hamamelidaceae, Moraceae, Rosaceae, Ulmus | ||
| eu-x.dd.mb.lhs | Eurythermic xeric drought-deciduous malacophyll low or high shrub | Artemisia, Asteraceae (excl.Artemisia), Brassicaceae, Euphorbiaceae, Fabaceae | ||
| tr-dt.lv | Tropical drought-tolerant liana or vine | Euphorbiaceae, Fabaceae, Oleaceae | ||
| tr-di.lv | Tropical drought-intolerant liana or vine | Apocynaceae, Fabaceae, Moraceae, Oleaceae, Ranunculaceae | ||
| te-dt.fb | Temperate drought-tolerant forb | Artemisia, Asteraceae (excl.Artemisia), Brassicaceae, Caryophyllaceae, Euphorbiaceae, Fabaceae, Lamiaceae, Polygonum, Rosaceae, Saxifragaceae, Scrophulariaceae | ||
| eu-dt.fb | Eurythermic drought-tolerant forb | Chenopodiaceae | ||
| g | Grass graminoid | Poaceae | ||
| lsuc | Leaf succulent | Chenopodiaceae | ||
| ssuc | Stem succulent | Euphorbiaceae | ||
Table S2 Main biome definitions in terms of plant functional type (Ni et al., 2010) |
| Biomes | Codes | PFTs |
|---|---|---|
| Cool evergreen needleleaf forest | COEG | eu.e.n.t, te-ft.cd.mb.t |
| Cool mixed forest | COMX | eu.e.n.t, te-fa.cd.mb.t, te-ft.cd.mb.t, te.cd.mb.lhs |
| Temperate deciduous broadleaf forest | TEDE | eu.e.n.t, te-fa.cd.mb.t, te-fi.cd.mb.t, te.cd.mb.lhs |
| Warm-temperate evergreen broadleaf and mixed forest | WTEM | wt.e.n.t, eu.e.n.t, wt.d.n.t, wt.e.sb.t, wt.e.mb.t, te-fi.cd.mb.t, wt.cd.mb.t, wt.e.sb.lhs, wt.e.mb.lhs, wt.cd.mb.lhs |
| Warm-temperate evergreen broadleaf forest | WTEG | wt.e.n.t, eu.e.n.t, wt.e.sb.t, wt.e.mb.t, wt.e.sb.lhs, wt.e.mb.lhs |
| Tropical semi-evergreen broadleaf forest | TRSE | wt.e.n.t, wt.d.n.t, wt.e.sb.t, wt.e.mb.t, tr.e.mb.t, tr-m.dd.mb.t, tr.e.mb.lhs, tr-dt.lv |
| Tropical evergreen broadleaf forest | TREG | wt.e.n.t, wt.d.n.t, wt.e.sb.t, wt.e.mb.t, tr.e.sb.t, tr.e.mb.t, tr.e.mb.lhs, tr-di.lv |
| Temperate xerophytic shrubland | TEXE | sl.t, dt.sl.lhs, te-dt.fb, g |
| Temperate grassland | TEGR | te-dt.fb, eu-dt.fb, g |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
China National Petroleum Corporation, 2000. Analysis and Identification on Spore-pollens. Beijing: National Petroleum and Chemical Industry Administration. (in Chinese)
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
Editorial Committee of China Vegetation Map CAS, 2001. 1:1,000,000 Scale Vegetation Distribution Map of China. Beijing: Science Press. (in Chinese)
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
IDoBM Center, 1983. Land Climate Data of China (1951-1980). Beijing: China Meteorological Press.
|
| [47] |
Institute of Botany CAS, 1982. Angiosperm Pollen Flora of Tropic and Subtropic China. Beijing: Science Press. (in Chinese)
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}