
Journal of Geographical Sciences >
Spatial distribution changes and habitat conservation of medicinal plant diversity in the Yinshan Mountains (China) under climate change
|
Zhao Zeyuan (2000‒), Master Candidate, specialized in protection and development and utilization of Chinese and Mongolian medicine resources. E-mail: zzy13763690405@163.com |
Received date: 2024-05-01
Accepted date: 2025-04-24
Online published: 2025-08-26
Supported by
The National Key Research and Development Program of China(2021YFE0190100)
Inner Mongolia Autonomous Region Mongolian Medicine Standardization Project(2023-[MB023])
The Earmarked Fund for CARS(CARS-21)
Medicinal plant diversity (MPD) is an indispensable part of global plant diversity, serving as the foundation for human survival by offering remedies and preventive measures against diseases. However, factors such as overexploitation, competition from invasive alien species, and climate change, threaten the habitats of medicinal plants, necessitating a comprehensive understanding of their spatial distribution and suitable habitats. We leveraged a decade of field survey data on medicinal plant distribution in the Yinshan Mountains, combined with spatial analysis, species distribution modeling, and the Carnegie Ames Stanford Approach (CASA) to explore the MPD spatial distribution and suitable habitats. Spatial analysis revealed that the central and eastern parts of Yinshan Mountains were the primary MPD hotspots, with no cold spots evident at various spatial scales. As the spatial scale decreased, previous non-significant regions transformed into hotspots, with instances where large-scale hotspots became insignificant. These findings offer valuable guidance for safeguarding and nurturing MPD across diverse spatial scales. In future climate change scenarios within the shared socioeconomic pathways (SSP), the habitat suitability for MPD in the Yinshan Mountains predominantly remains concentrated in the central and eastern regions. Notably, areas with high net primary productivity (NPP) values and abundant vegetation coverage align closely with MPD habitat suitability areas, potentially contributing to the region’s rich MPD.
ZHAO Zeyuan , BI Yaqiong , WEI Xinxin , CHEN Yuan , ZHANG Ru , GUO Jingxia , ZHANG Mingxu , ZHANG Xiaobo , LI Minhui . Spatial distribution changes and habitat conservation of medicinal plant diversity in the Yinshan Mountains (China) under climate change[J]. Journal of Geographical Sciences, 2025 , 35(7) : 1479 -1496 . DOI: 10.1007/s11442-025-2380-x
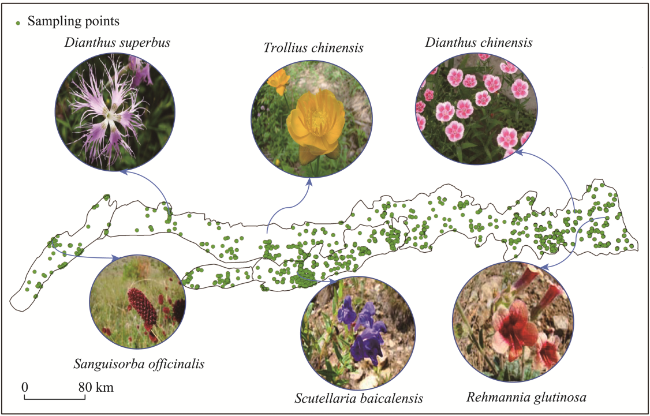
Figure 1 Sampling points used to study medicinal plant distributions in the Yinshan Mountains |
Supplementary Table 1 The environment variables information |
| Abbreviation | Environment variables | Unit |
|---|---|---|
| Bio_1 | Annual mean temperature | ℃ |
| Bio_2 | Mean diurnal range (Mean of monthly (max temp-min temp)) | ℃ |
| Bio_3 | Isothermality (Bio_2/Bio_7) (×100) | - |
| Bio_4 | Temperature seasonality (standard deviation × 100) | - |
| Bio_5 | Max temperature of warmest month | ℃ |
| Bio_6 | Min temperature of coldest month | ℃ |
| Bio_7 | Temperature annual range (Bio_5-Bio_6) | ℃ |
| Bio_8 | Mean temperature of wettest quarter | ℃ |
| Bio_9 | Mean temperature of driest quarter | ℃ |
| Bio_10 | Mean temperature of warmest quarter | ℃ |
| Bio_11 | Mean temperature of coldest quarter | ℃ |
| Bio_12 | Annual precipitation | mm |
| Bio_13 | Precipitation of wettest month | mm |
| Bio_14 | Precipitation of driest month | mm |
| Bio_15 | Precipitation seasonality (Coefficient of Variation) | - |
| Bio_16 | Precipitation of wettest quarter | mm |
| Bio_17 | Precipitation of driest quarter | mm |
| Bio_18 | Precipitation of warmest quarter | mm |
| Bio_19 | Precipitation of coldest quarter | mm |
| Elv | Elevation | m |
Supplementary Table 2 List of plants from three sources in MaxEnt analysis |
| Species | Source | Species | Source | Species | Source | |||
|---|---|---|---|---|---|---|---|---|
| Agrimonia pilosa | more than 60 points | Euphorbia esula | 10-30 points | Saussurea japonica | 10-30 points | |||
| Artemisia scoparia | more than 60 points | Fagopyrum esculentum | 10-30 points | Saussurea nivea | 10-30 points | |||
| Artemisia stechmanniana | more than 60 points | Fagopyrum tataricum | 10-30 points | Scabiosa comosa | 10-30 points | |||
| Bistorta officinalis | more than 60 points | Fallopia convolvulus | 10-30 points | Schizonepeta multifida | 10-30 points | |||
| Bupleurum chinense | more than 60 points | Filifolium sibiricum | 10-30 points | Schizonepeta tenuifolia | 10-30 points | |||
| Bupleurum scorzonerifolium | more than 60 points | Gentiana macrophylla | 10-30 points | Scorzonera sinensis | 10-30 points | |||
| Delphinium grandiflorum | more than 60 points | Geranium wilfordii | 10-30 points | Scutellaria scordifolia | 10-30 points | |||
| Dianthus chinensis | more than 60 points | Geum aleppicum | 10-30 points | Scutellaria viscidula | 10-30 points | |||
| Echinops sphaerocephalus | more than 60 points | Glycyrrhiza uralensis | 10-30 points | Silene aprica | 10-30 points | |||
| Gentiana dahurica | more than 60 points | Grubovia dasyphylla | 10-30 points | Silene repens | 10-30 points | |||
| Medicago ruthenica | more than 60 points | Halenia corniculata | 10-30 points | Sonchus wightianus | 10-30 points | |||
| Polygala tenuifolia | more than 60 points | Haplophyllum dauricum | 10-30 points | Sphallerocarpus gracilis | 10-30 points | |||
| Potentilla chinensis | more than 60 points | Hedysarum brachypterum | 10-30 points | Spiraea pubescens | 10-30 points | |||
| Sanguisorba officinalis | more than 60 points | Hibiscus trionum | 10-30 points | Spiraea salicifolia | 10-30 points | |||
| Saposhnikovia divaricata | more than 60 points | Hippophae rhamnoides | 10-30 points | Stellaria dichotoma | 10-30 points | |||
| Scutellaria baicalensis | more than 60 points | Hylotelephium malacophyllum | 10-30 points | Takhtajaniantha austriaca | 10-30 points | |||
| Stellera chamaejasme | more than 60 points | Hyoscyamus niger | 10-30 points | Teloxys aristata | 10-30 points | |||
| Taraxacum mongolicum | more than 60 points | Imperata cylindrica | 10-30 points | Thalictrum aquilegiifolium | 10-30 points | |||
| Thalictrum petaloideum | more than 60 points | Incarvillea sinensis | 10-30 points | Thalictrum squarrosum | 10-30 points | |||
| Ulmus pumila | more than 60 points | Inula britannica | 10-30 points | Thermopsis lanceolata | 10-30 points | |||
| Aconitum barbatum | 10-30 points | Inula japonica | 10-30 points | Tournefortia sibirica | 10-30 points | |||
| Aconitum kusnezoffii | 10-30 points | Iris dichotoma | 10-30 points | Tribulus terrestris | 10-30 points | |||
| Adenophora capillaris | 10-30 points | Iris lactea | 10-30 points | Trifolium lupinaster | 10-30 points | |||
| Adenophora polyantha | 10-30 points | Ixeris chinensis | 10-30 points | Triglochin palustris | 10-30 points | |||
| Agropyron cristatum | 10-30 points | Ixeris polycephala | 10-30 points | Trollius chinensis | 10-30 points | |||
| Allium condensatum | 10-30 points | Klasea centauroides | 10-30 points | Ulmus macrocarpa | 10-30 points | |||
| Allium mongolicum | 10-30 points | Knorringia sibirica | 10-30 points | Urtica cannabina | 10-30 points | |||
| Allium senescens | 10-30 points | Lactuca tatarica | 10-30 points | Valeriana officinalis | 10-30 points | |||
| Allium tenuissimum | 10-30 points | Lagopsis supina | 10-30 points | Veronica anagallis-aquatica | 10-30 points | |||
| Amaranthus retroflexus | 10-30 points | Lappula myosotis | 10-30 points | Vicia cracca | 10-30 points | |||
| Amethystea caerulea | 10-30 points | Larix gmelinii | 10-30 points | Viola variegata | 10-30 points | |||
| Argentina anserina | 10-30 points | Leibnitzia anandria | 10-30 points | Xanthium strumarium | 10-30 points | |||
| Artemisia annua | 10-30 points | Leonurus japonicus | 10-30 points | Actaea dahurica | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Artemisia caruifolia | 10-30 points | Leonurus sibiricus | 10-30 points | Adenophora gmelinii | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Artemisia eriopoda | 10-30 points | Leptopyrum fumarioides | 10-30 points | Adenophora stenanthina | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Artemisia frigida | 10-30 points | Lespedeza cuneata | 10-30 points | Adenophora tetraphylla | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Artemisia gmelinii | 10-30 points | Lespedeza davurica | 10-30 points | Agropyron mongolicum | List of National Key Protected Wild Plants in China | |||
| Artemisia mongolica | 10-30 points | Lespedeza juncea | 10-30 points | Anemarrhena asphodeloides | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Artemisia sacrorum | 10-30 points | Lespedeza potaninii | 10-30 points | Anemone sylvestris | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Artemisia tanacetifolia | 10-30 points | Lilium pumilum | 10-30 points | Astragalus membranaceus | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Asparagus dauricus | 10-30 points | Limonium bicolor | 10-30 points | Atraphaxis bracteata | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Aster hispidus | 10-30 points | Linaria vulgaris | 10-30 points | Berberis caroli | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Aster tataricus | 10-30 points | Linum stelleroides | 10-30 points | Bistorta officinalis | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Astragalus chinensis | 10-30 points | Linum usitatissimum | 10-30 points | Bupleurum sibiricum | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Astragalus dahuricus | 10-30 points | Malva verticillata | 10-30 points | Campanula punctata | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Astragalus scaberrimus | 10-30 points | Medicago lupulina | 10-30 points | Caryopteris mongholica | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Atractylodes chinensis | 10-30 points | Medicago sativa | 10-30 points | Clematis fruticosa | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Bassia scoparia | 10-30 points | Melilotus albus | 10-30 points | Cnidium monnieri | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Belamcanda chinensis | 10-30 points | Melilotus suaveolens | 10-30 points | Cornus officinalis | Wild herbal resources protection management regulations | |||
| Betula platyphylla | 10-30 points | Mentha canadensis | 10-30 points | Dasiphora glabra | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Bidens parviflora | 10-30 points | Neopallasia pectinata | 10-30 points | Eleutherococcus senticosus | Wild herbal resources protection management regulations | |||
| Bidens pilosa | 10-30 points | Odontites vulgaris | 10-30 points | Elsholtzia ciliata | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Bothriospermum kusnetzowii | 10-30 points | Olgaea lomonossowii | 10-30 points | Ephedra equisetina | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Bupleurum mithii | 10-30 points | Oreomecon nudicaulis | 10-30 points | Ephedra monosperma | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Calamagrostis epigeios | 10-30 points | Orobanche pycnostachya | 10-30 points | Ephedra sinica | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Callistephus chinensis | 10-30 points | Orostachys fimbriata | 10-30 points | Equisetum arvense | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Cannabis sativa | 10-30 points | Ostryopsis davidiana | 10-30 points | Eucommia ulmoides | Wild herbal resources protection management regulations | |||
| Caragana microphylla | 10-30 points | Oxybasis glauca | 10-30 points | Forsythia suspensa | Wild herbal resources protection management regulations | |||
| Caragana sinica | 10-30 points | Oxytropis coerulea | 10-30 points | Gentiana macrophylla | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region, Wild herbal resources protection management regulations | |||
| Carduus crispus | 10-30 points | Paeonia lactiflora | 10-30 points | Ginkgo biloba | List of National Key Protected Wild Plants in China | |||
| Carduus nutans | 10-30 points | Parthenocissus tricuspidata | 10-30 points | Glycyrrhiza uralensis | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region, Wild herbal resources protection management regulations | |||
| Carum buriaticum | 10-30 points | Patrinia rupestris | 10-30 points | Haplophyllum tragacanthoides | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Caryopteris mongholica | 10-30 points | Patrinia scabiosifolia | 10-30 points | Hemerocallis minor | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Catolobus pendulus | 10-30 points | Pedicularis resupinata | 10-30 points | Leonurus sibiricus | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Chelidonium majus | 10-30 points | Pedicularis striata | 10-30 points | Lilium pumilum | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Chenopodium ficifolium | 10-30 points | Persicaria lapathifolia | 10-30 points | Limonium bicolor | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Chloris virgata | 10-30 points | Phedimus aizoon | 10-30 points | Malaxis monophyllos | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Chrysanthemum chanetii | 10-30 points | Phlomoides umbrosa | 10-30 points | Mentha canadensis | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Cirsium arvense | 10-30 points | Phragmites australis | 10-30 points | Olgaea lomonossowii | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Clematis brevicaudata | 10-30 points | Polygala sibirica | 10-30 points | Orobanche pycnostachya | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Clematis hexapetala | 10-30 points | Polygonatum odoratum | 10-30 points | Paeonia lactiflora | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Clematis intricata | 10-30 points | Polygonatum sibiricum | 10-30 points | Phedimus aizoon | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Convolvulus ammannii | 10-30 points | Populus simonii | 10-30 points | Platycodon grandiflorus | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Convolvulus tragacanthoides | 10-30 points | Portulaca oleracea | 10-30 points | Polygala sibirica | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Cuscuta chinensis | 10-30 points | Potentilla betonicifolia | 10-30 points | Polygala tenuifolia | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region, Wild herbal resources protection management regulations | |||
| Cymbaria dahurica | 10-30 points | Potentilla bifurca | 10-30 points | Polygonatum odoratum | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Cynanchum chinense | 10-30 points | Potentilla multicaulis | 10-30 points | Polygonatum sibiricum | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Dasiphora fruticosa | 10-30 points | Potentilla tanacetifolia | 10-30 points | Pulsatilla chinensis | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Descurainia sophia | 10-30 points | Prunus armeniaca | 10-30 points | Ranunculus japonicus | the Convention on International Trade in Endangered Species of Wild Fauna and Flora | |||
| Dianthus superbus | 10-30 points | Prunus sibirica | 10-30 points | Rheum franzenbachii | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Diarthron linifolium | 10-30 points | Prunus triloba | 10-30 points | Saposhnikovia divaricata | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region, Wild herbal resources protection management regulations | |||
| Dontostemon dentatus | 10-30 points | Pseudolysimachion linariifolium | 10-30 points | Schisandra chinensis | Wild herbal resources protection management regulations | |||
| Dracocephalum rupestre | 10-30 points | Pulsatilla chinensis | 10-30 points | Scutellaria baicalensis | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region, Wild herbal resources protection management regulations | |||
| Echinochloa crusgalli | 10-30 points | Pulsatilla turczaninovii | 10-30 points | Sophora alopecuroides | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Elsholtzia ciliata | 10-30 points | Rehmannia glutinosa | 10-30 points | Sophora flavescens | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Elymus dahuricus | 10-30 points | Rosa xanthina | 10-30 points | Stellaria dichotoma | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Equisetum arvense | 10-30 points | Rubia cordifolia | 10-30 points | Trollius chinensis | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Equisetum ramosissimum | 10-30 points | Rubus saxatilis | 10-30 points | Vincetoxicum pycnostelma | List of Key Protected Wild Plants in Inner Mongolia Autonomous Region | |||
| Eremogone juncea | 10-30 points | Salsola komarovii | 10-30 points | |||||
| Eritrichium rupestre | 10-30 points | Saussurea amara | 10-30 points | |||||
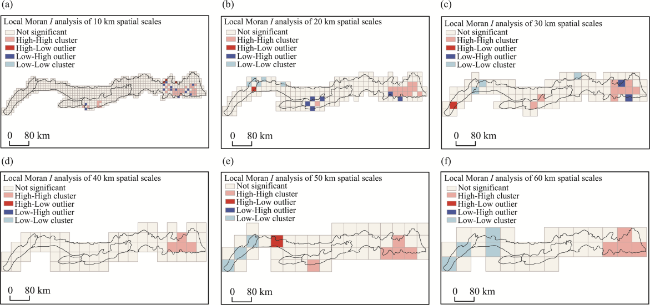
Supplementary Table 3 Results of spatial autocorrelation analysis at different spatial scales |
| Grid size | Moran’s I | Z | P |
|---|---|---|---|
| 10 | 0.324690 | 11.288268 | 0.000000 |
| 20 | 0.360309 | 6.773821 | 0.000000 |
| 30 | 0.480294 | 6.258639 | 0.000000 |
| 40 | 0.416629 | 4.544818 | 0.000005 |
| 50 | 0.427541 | 4.033750 | 0.000055 |
| 60 | 0.535239 | 4.098584 | 0.000042 |
Figure 2 Local Moran’s I analysis of the Yinshan Mountains at different scales: (a) 10 km; (b) 20 km; (c) 30 km; (d) 40 km; (e) 50 km; (f) 60 km |
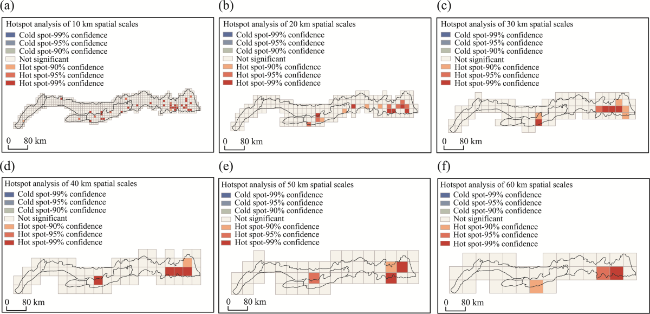
Figure 3 Hotspot analysis of the Yinshan Mountains at different scales: (a) 10 km; (b) 20 km; (c) 30 km; (d) 40 km; (e) 50 km; (f) 60 km |
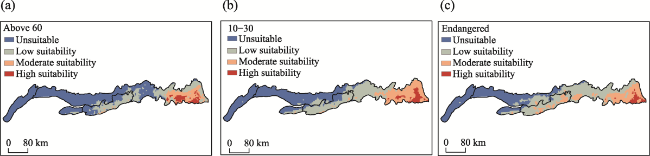
Figure 4 Habitat suitability distribution of MPD in the Yinshan Mountains: (a) Widely distributed medicinal plants; (b) Rare distribution of medicinal plants; (c) Endangered medicinal plants |
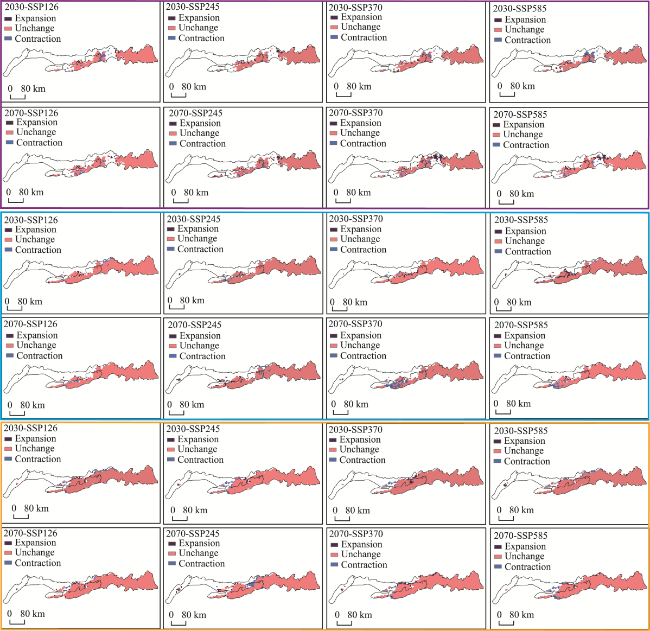
Figure 5 Habitat suitability distribution of MPD in the Yinshan Mountains under future climate (Purple represents expansion, pink represents unchanged, and blue represents contraction.) |
Supplementary Table 4 Potential distribution changes under future climatic conditions (km2) |
| Species | Period Scenario | Range expansion | No change | Range contraction | |
|---|---|---|---|---|---|
| Widely distributed medicinal plants (Above 60) | 2021-2040 | SSP126 | 754.89 | 18760.29 | 1062.43 |
| SSP245 | 866.72 | 19235.59 | 587.13 | ||
| SSP370 | 698.97 | 18983.96 | 838.76 | ||
| SSP585 | 698.97 | 17949.49 | 1873.23 | ||
| 2061-2080 | SSP126 | 615.09 | 18872.12 | 950.60 | |
| SSP245 | 1090.39 | 18592.54 | 1230.18 | ||
| SSP370 | 1845.27 | 18592.54 | 1230.18 | ||
| SSP585 | 1677.52 | 18956.00 | 866.72 | ||
| Rare distribution of medicinal plants (10-30) | 2021-2040 | SSP126 | 335.50 | 26225.26 | 810.80 |
| SSP245 | 279.59 | 26057.51 | 978.55 | ||
| SSP370 | 671.01 | 26504.85 | 531.22 | ||
| SSP585 | 1369.98 | 26700.56 | 335.50 | ||
| 2061-2080 | SSP126 | 195.71 | 26281.18 | 754.88 | |
| SSP245 | 1090.39 | 26113.43 | 922.64 | ||
| SSP370 | 111.83 | 24379.99 | 2656.08 | ||
| SSP585 | 335.50 | 25694.05 | 1342.02 | ||
| Endangered medicinal plants (Endangered) | 2021-2040 | SSP126 | 698.97 | 30083.56 | 1118.35 |
| SSP245 | 726.93 | 30111.52 | 1090.39 | ||
| SSP370 | 810.80 | 30307.23 | 894.68 | ||
| SSP585 | 643.05 | 30251.32 | 950.60 | ||
| 2061-2080 | SSP126 | 726.93 | 30754.57 | 447.34 | |
| SSP245 | 950.60 | 29636.22 | 1565.69 | ||
| SSP370 | 1034.47 | 30335.19 | 866.72 | ||
| SSP585 | 726.93 | 29859.89 | 1342.02 | ||
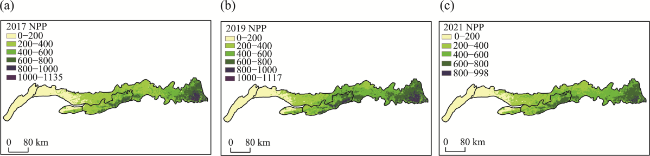
Figure 6 NPP values in the Yinshan Mountains: (a) 2017; (b) 2019; (c) 2021 |
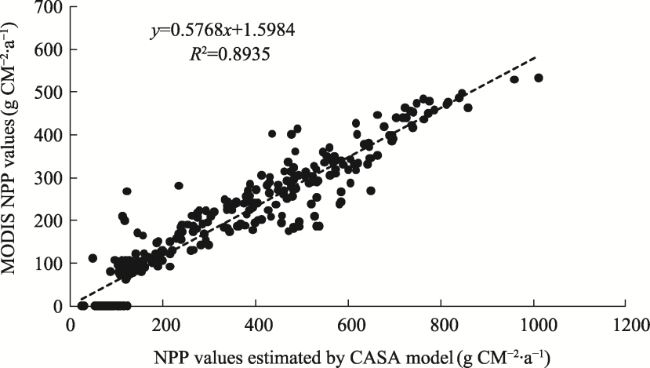
Figure 7 Correlation between MODIS NPP values and NPP values estimated by the CASA model |
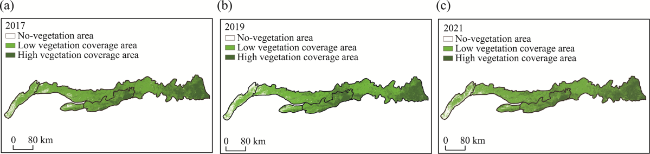
Figure 8 NDVI values in the Yinshan Mountains: (a) 2017; (b) 2019; (c) 2021 |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
China Medicinal Materials Company, 1994. China Traditional Chinese Medicine Resource Series, China Traditional Chinese Medicine Resource Series Summary. Beijing: Science Press. (in Chinese)
|
| [7] |
Chinese Academy of Sciences Inner Mongolia and Ningxia Comprehensive Expedition Team, 1985. Inner Mongolia Vegetation. Beijing: Science Press. (in Chinese)
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}