
Journal of Geographical Sciences >
Elevation response of above-ground net primary productivity for Picea crassifolia to climate change in Qilian Mountains of Northwest China based on tree rings
|
Wu Xuan (1996-), specialized in dendrochronology. E-mail: 15002550410@163.com. |
Received date: 2022-07-04
Accepted date: 2023-07-19
Online published: 2024-01-08
Supported by
The CAS “Light of West China” Program(2020XBZG-XBQNXZ-A)
Cultivation Program of 2022 Major Scientific Research Project of Northwest Normal University(WNU-LKZD2022-04)
National Natural Science Foundation of Gansu(20JR10RA093)
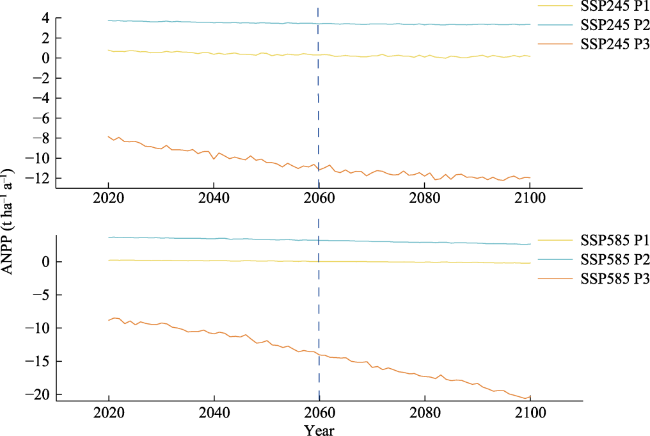
Current ecosystem models used to simulate global terrestrial carbon balance generally suggest that terrestrial landscapes are stable and mature, but terrestrial net primary productivity (NPP) data estimated without accounting for disturbances in species composition, environment, structure, and ecological characteristics will reduce the accuracy of the global carbon budget. Therefore, the steady-state assumption and neglect of elevation-related changes in forest NPP is a concern. The Qilian Mountains are located in continental climate zone, and vegetation is highly sensitive to climate change. We quantified aboveground biomass (AGB) and aboveground net primary productivity (ANPP) sequences at three elevations using field-collected tree rings of Picea crassifolia in Qilian Mountains of Northwest China. The results showed that (1) There were significant differences between AGB and ANPP at the three elevations, and the growth rate of AGB was the highest at the low elevation (55.99 t ha-1 10a-1). (2) There are differences in the response relationship between the ANPP and climate factors at the three elevations, and drought stress is the main climate signal affecting the change of ANPP. (3) Under the future climate scenario, drought stress intensifies, and the predicted decline trend of ANPP at the three elevations from mid-century to the end of this century is -0.025 t ha-1 10a-1, respectively; -0.022 t ha-1 10a-1; At -0.246 t ha-1 10a-1, the level of forest productivity was significantly degraded. The results reveal the elevation gradient differences in forest productivity levels and provide key information for studying the carbon sink potential of boreal forests.
WU Xuan , JIAO Liang , DU Dashi , XUE Ruhong , WEI Mengyuan , ZHANG Peng . Elevation response of above-ground net primary productivity for Picea crassifolia to climate change in Qilian Mountains of Northwest China based on tree rings[J]. Journal of Geographical Sciences, 2024 , 34(1) : 146 -164 . DOI: 10.1007/s11442-024-2199-x
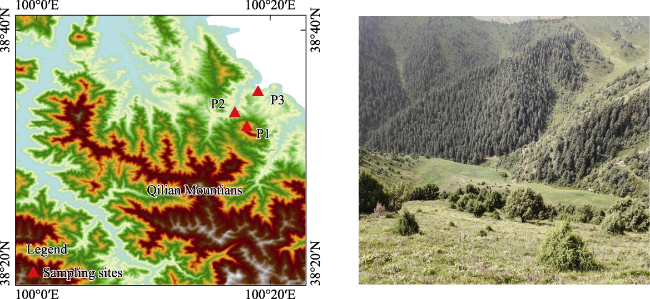
Figure 1 Distribution of the sampling sites |
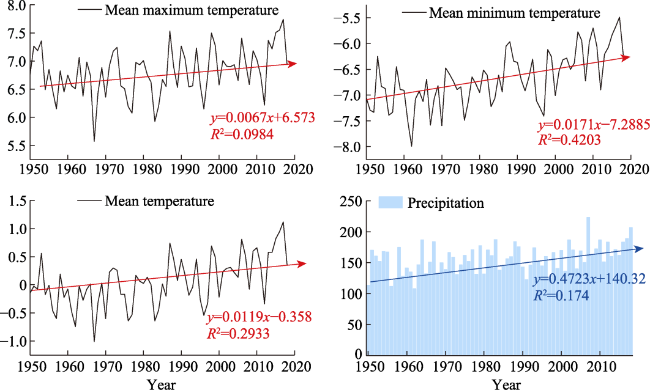
Figure 2 Interannual variation of meteorological factors (mean maximum temperature, mean temperature, mean minimum temperature and total precipitation) |
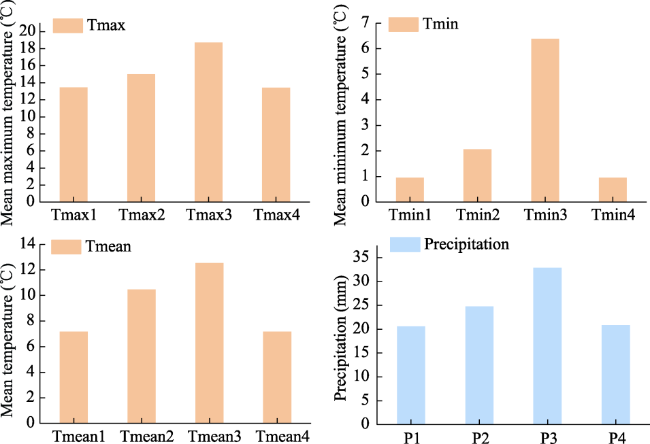
Figure 3 Tmax, Tmin, Tmean and precipitation at different stages of the growing season in the Qilian Mountains of Northwest China. 1, 2, 3 and 4 represent the end of the previous growing season, the beginning of the current growing season, the middle of the current growing season, and the end of the current growing season. |
Table 1 Information about sampling sites of the Qinghai spruce at the three elevations |
| Site | Elevation (m) | Latitude (°N) | Longitude (°E) | Slope (°) | CC (%) | TD (m) | DBH (cm) | TH (m) | CW (m) |
|---|---|---|---|---|---|---|---|---|---|
| P1 | 3300 | 38.32 | 100.18 | 30 | 40.4 | 6.0 | 40.0 | 20.3 | 4.1 |
| P2 | 2850 | 38.33 | 100.17 | 17 | 50.2 | 5.2 | 30.8 | 14.5 | 4.0 |
| P3 | 2585 | 38.35 | 100.19 | 19 | 45.3 | 7.2 | 31.1 | 16.3 | 4.2 |
P is the sampling site, P1: High elevation, P2: Middle elevation, P3: Low elevation. CC (%): canopy coverage, TD: tree distance, DBH: diameter at breast height, TH: tree height, CW: crown width. |
Table 2 Dendrochronological characteristics of chronologies for the Qinghai spruce at the three elevations |
| Sites | P1 | P2 | P3 |
|---|---|---|---|
| Core/tree | 49/25 | 48/24 | 48/24 |
| Time periods | 1837-2019 | 1819-2019 | 1902-2019 |
| MS | 0.224 | 0.280 | 0.307 |
| SD | 0.220 | 0.338 | 0.326 |
| ACI | 0.637 | 0.542 | 0.479 |
| R | 0.258 | 0.532 | 0.671 |
| R1 | 0.475 | 0.734 | 0.843 |
| R2 | 0.253 | 0.528 | 0.660 |
| PC1 | 0.303 | 0.563 | 0.703 |
| SNR | 23.259 | 86.353 | 63.084 |
| EPS | 0.959 | 0.989 | 0.984 |
P is the sampling site, P1: High elevation, P2: Middle elevation, P3: Low elevation. |
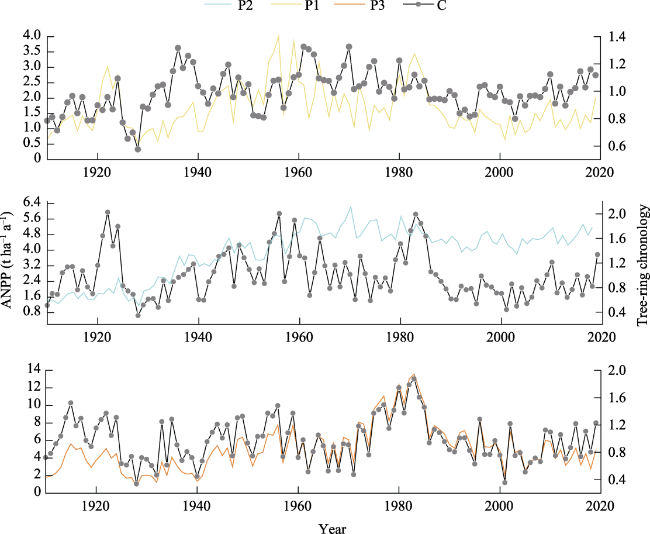
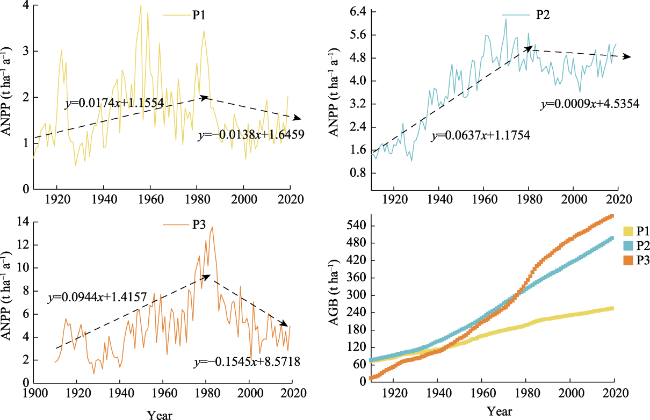
Figure 4 Interannual variations of ANPP and tree-ring chronologies at the three elevations |
Figure 5 Interannual variation of ANPP and AGB at the three elevations |
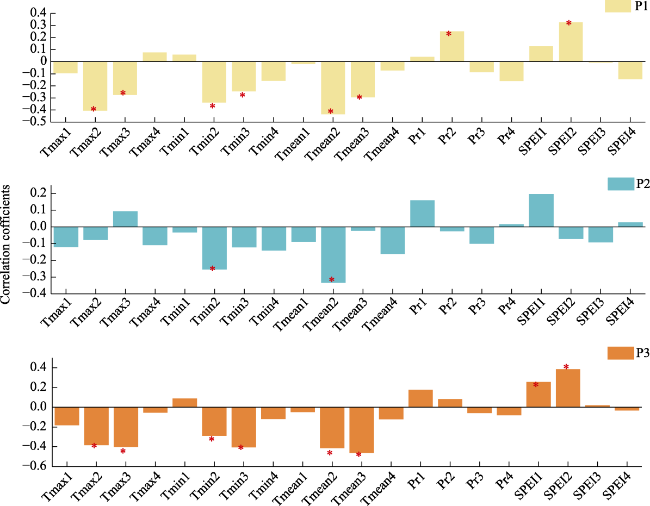
Figure 6 Correlation between climate factors and ANPP from 1950 to 2018 (max: mean maximum temperature, min: mean minimum temperature, mean: mean temperature, Pr: precipitation and SPEI) at the three elevations. 1, 2, 3 and 4 represent the end of the previous growing season, the beginning of the current growing season, the middle of the current growing season, and the end of the current growing season. |
Table 3 Integrated characteristic values of regression models at the three elevations |
| Independent variable | R² | Adjusted R² | Std. error of the estimate | F | P | |
|---|---|---|---|---|---|---|
| P1 | Tmean2 | 0.190 | 0.178 | 0.691 | 15.712 | 0.000 |
| Tmean2 Pr2 | 0.244 | 0.221 | 0.673 | 10.670 | 0.000 | |
| P2 | Tmean2 | 0.110 | 0.097 | 0.521 | 8.314 | 0.005 |
| P3 | Tmean3 | 0.209 | 0.197 | 2.355 | 17.724 | 0.000 |
| Tmean3 SPEI2 | 0.310 | 0.289 | 2.216 | 14.819 | 0.000 | |
| Tmean3 SPEI2 SPEI1 | 0.374 | 0.345 | 2.127 | 12.956 | 0.000 | |
| Tmean3 SPEI2 SPEI1 Tmin2 | 0.428 | 0.392 | 2.050 | 11.953 | 0.000 |
Table 4 Regression schemes and eigenvalues of ANPP at the three elevations |
| Number | Model | B | Std. error | Standardized coefficients | t | Sig. | |
|---|---|---|---|---|---|---|---|
P1 | 1 | Constant | 6.413 | 1.170 | 5.48 | 0.000 | |
| Tmean2 | -0.443 | 0.112 | -0.436 | -3.964 | 0.000 | ||
| 2 | Constant | 5.967 | 1.157 | 5.156 | 0.000 | ||
| Tmean2 | -0.433 | 0.109 | -0.426 | -3.980 | 0.000 | ||
| Pr2 | 0.018 | 0.008 | 0.233 | 2.179 | 0.033 | ||
| P2 | 1 | Constant | 7.150 | 0.883 | 8.079 | 0.000 | |
| Tmean2 | -0.243 | 0.084 | -0.332 | -2.883 | 0.005 | ||
| P3 | 1 | Constant | 29.801 | 5.628 | 5.295 | 0.000 | |
| Tmean3 | -1.893 | 0.450 | -0.457 | -4.210 | 0.000 | ||
| 2 | Constant | 27.326 | 5.357 | 5.101 | 0.000 | ||
| Tmean3 | -1.695 | 0.428 | -0.410 | -3.962 | 0.000 | ||
| SPEI2 | 1.004 | 0.323 | 0.321 | 3.103 | 0.003 | ||
| 3 | Constant | 26.847 | 5.144 | 5.219 | 0.000 | ||
| Tmean3 | -1.658 | 0.411 | -0.401 | -4.035 | 0.000 | ||
| SPEI2 | 1.039 | 0.311 | 0.332 | 3.345 | 0.001 | ||
| SPEI1 | 0.561 | 0.217 | 0.254 | 2.584 | 0.012 | ||
| 4 | Constant | 24.347 | 5.062 | 4.809 | 0.000 | ||
| Tmean3 | -1.306 | 0.421 | -0.316 | -3.101 | 0.003 | ||
| SPEI2 | 1.154 | 0.303 | 0.369 | 3.807 | 0.000 | ||
| SPEI1 | 0.628 | 0.211 | 0.284 | 2.973 | 0.004 | ||
| Tmin2 | -0.924 | 0.378 | -0.248 | -2.443 | 0.017 |
P is the sampling site, P1: High elevation, P2: Middle elevation, P3: Low elevation. Tmax: mean maximum temperature, Tmin: mean minimum temperature, Tmean: mean temperature, Pr: precipitation at the three elevations. 1, 2, 3 and 4 represent the end of the previous growing season, the beginning of the current growing season, the middle of the current growing season, and the end of the current growing season. |
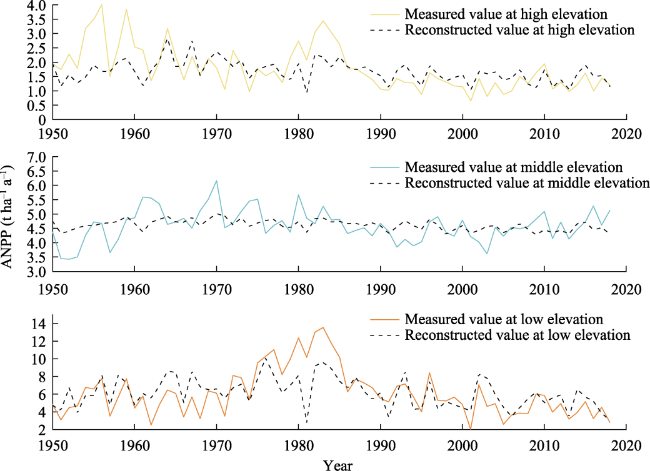
Figure 7 Reconstructed value of ANPP fitted by regression scheme was compared with the actual value ANPP |
Figure 8 Interannual variation of ANPP at the three elevations under two future climate scenarios |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}