
Journal of Geographical Sciences >
High leaf area index expands the contrasting effect of climate warming on Western Siberia taiga forests activity before and after 2000
|
Sun Han (1988-), Associate Professor, specialized in climatic change, temperature, timberline, net primary productivity and ecology. E-mail: sunhan@bjfu.edu.cn |
Received date: 2023-05-02
Accepted date: 2023-11-07
Online published: 2024-01-08
Supported by
The Third Xinjiang Scientific Expedition Program(2021xjkk0603)
The Third Xinjiang Scientific Expedition Program(2022xjkk1205)
National Natural Science Foundation of China(32201258)
National Natural Science Foundation of China(32271652)
The taiga vegetation in Western Siberia has been seriously threatened by climate warming in recent decades. However, how vegetation in different growing states and climate conditions responds to climate changes differently is still unclear. Here we explore the vegetation activity trends in Western Siberia taiga forests using the annual rate of change in leaf area index (LAI) during 1982-2018 so as to answer two questions: (1) how did climate warming affect taiga vegetation activity in the recent last decades? (2) Did the growing state of taiga forest affect its response to climate warming? Our results revealed that climate warming promoted taiga vegetation activity in Western Siberia before 2000. However, continuous warming caused excessive evapotranspiration and led to decreased vegetation activity after 2000. Moreover, the intensity of vegetation growth response to warming was positively related to canopy height and LAI, indicating that both the positive and negative effects of warming were more significant in taiga forests in better growing state. Since these forests generally have higher productivity and play more important roles in ecosystem functioning (e.g., carbon sink and biodiversity conservation), our results highlight their vulnerability to future climate change that need more research attention.
SUN Han , WANG Xiangping . High leaf area index expands the contrasting effect of climate warming on Western Siberia taiga forests activity before and after 2000[J]. Journal of Geographical Sciences, 2024 , 34(1) : 131 -145 . DOI: 10.1007/s11442-024-2198-y
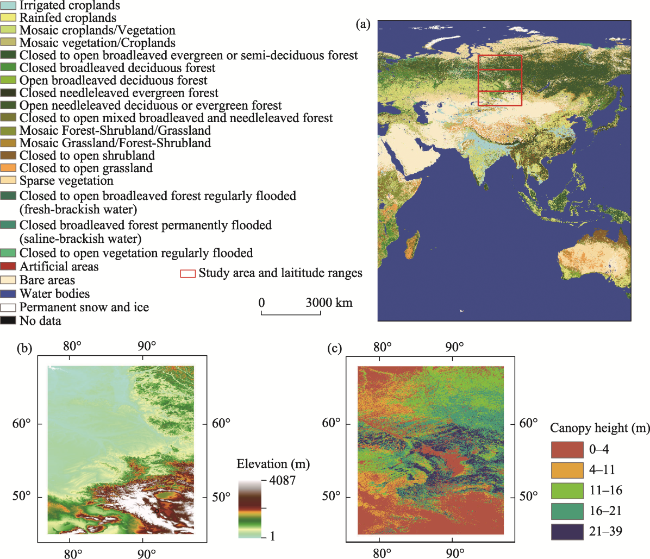
Figure 1 (a) Location of the study region in Eurasia. The land cover types were determined according to the global land cover data at a resolution of 300 m (GlobCover 2009 v2.3) which was produced via global collaborative efforts (Arino et al., 2008). (b) Topography of the study region. The elevation data was derived from the Shuttle Radar Topography Mission (SRTM) digital elevation datasets at a resolution of 90 m (Jarvis et al., 2008). (c) Canopy height patterns across the study region. The forest height data were extracted from a global forest height map, which was estimated from satellite Lidar (light detection and ranging) data (Simard et al., 2011). |
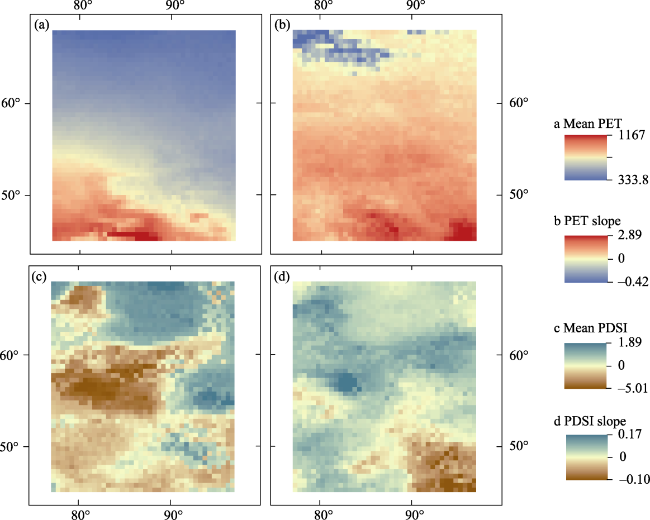
Figure 2 Spatial patterns of climatic conditions and climate change trends for taiga forests from 1982 to 2018. (a and c) Mean annual potential evapotranspiration (PET) and Palmer drought severity index (PSDI) from 1982 to 2018. (b and d) Slope of PET and PDSI change during 1982-2018, respectively. |
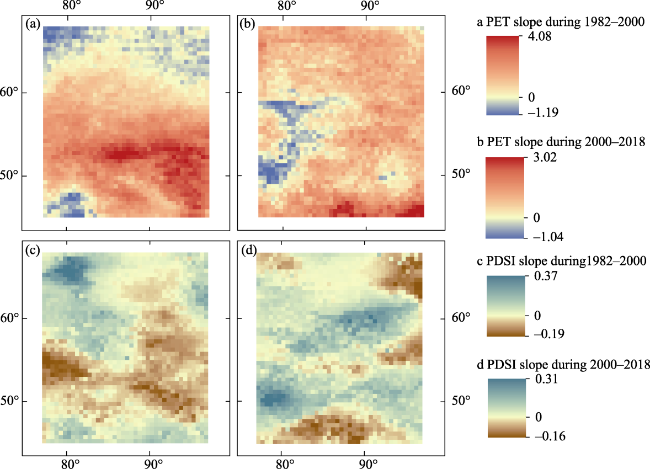
Figure 3 Spatial patterns of climate change trends before and after 2000. (a and c) Slope of potential evapotranspiration (PET) and Palmer drought severity index (PSDI) during 1982-2000. (b and d) PET and PDSI slope during 2000-2018, respectively. |
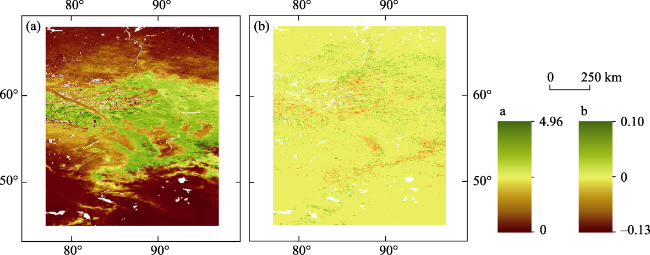
Figure 4 Spatial patterns of (a) multi-decade mean leaf area index (LAI), and (b) annual changing rate (slope) of LAI during 1982-2018 |
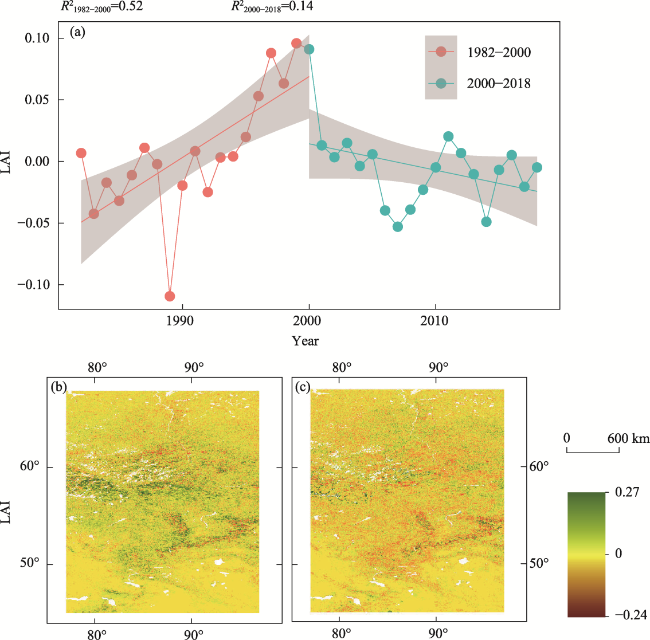
Figure 5 Contrasting temporal trends of leaf area index (LAI) before and after 2000. (a) Interannual variations in the mean LAI across the study area. R21982-2000 and R22000-2018 represent the R2 of the fitted line during 1982-2000 and 2000-2018, respectively. (b) and (c), spatial patterns of LAI slopes during 1982-2000 and 2000-2018, respectively. |
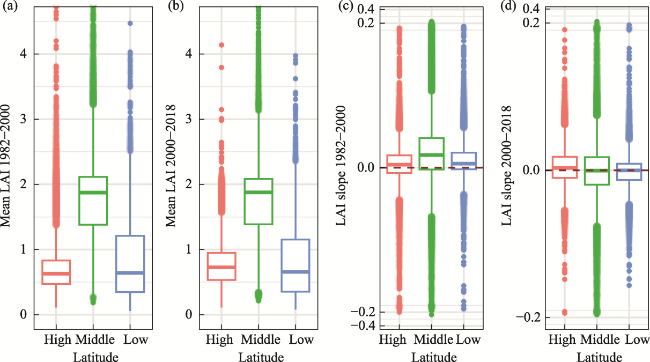
Figure 6 (a and b) Multi-year mean leaf area index (LAI) and (c and d) LAI changing rate (slope) before and after 2000 in the low, middle and high latitudinal zones of the study area |
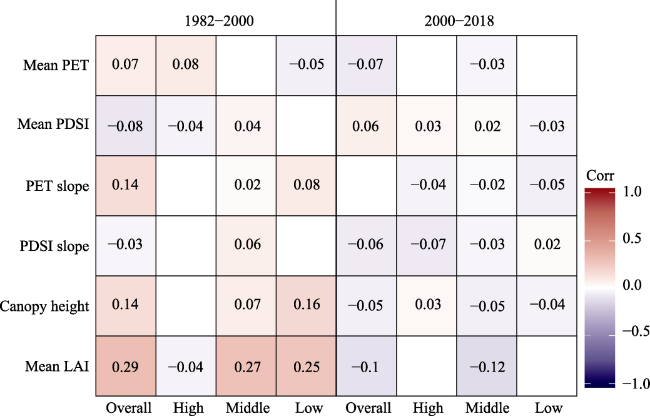
Figure 7 Correlations of climate and biotic factors with LAI slope before and after 2000, for the study area (overall) and for the high, middle and low latitudinal zones. Non-significant correlations (p > 0.05) were not shown. |
Table 1 Standardized relative importance of predictors in the random forest models to explain the LAI slope during 1982-2018 for the study area (overall) and for the high, middle, and south latitudinal zones. Each importance value was calculated in proportion to the maximum importance observed, and thus the highest importance was 1. More important variables (> 0.5) are boldfaced. |
| Overall | High | Middle | Low | |
|---|---|---|---|---|
| Mean PET | 0.48 | 0.40 | 0.39 | 0.65 |
| PET slope | 0.41 | 0.43 | 0.23 | 0.60 |
| Mean PDSI | 1.00 | 0.25 | 0.52 | 0.69 |
| PDSI slope | 0.89 | 0.47 | 0.41 | 0.61 |
| Canopy height | 0.36 | 0.46 | 0.56 | 0.35 |
| Mean LAI | 0.79 | 1.00 | 1.00 | 1.00 |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
IPCC, 2021. Climate Change 2021: The Physical Science Basis. IPCC: Switzerland.
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
University of East Anglia Climatic Research Unit, Harris I C,
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}