
Journal of Geographical Sciences >
Risk posed to vegetation net primary productivity by drought on the Mongolian Plateau
|
Ren Jinyuan (1995-), PhD Candidate, E-mail: renjiny1217@163.com |
Received date: 2023-03-14
Accepted date: 2023-07-03
Online published: 2023-11-15
Supported by
Natural Science Foundation of Inner Mongolia(2023MS04001)
National Natural Science Foundation of China(42061070)
National Natural Science Foundation of China(42261144746)
Program for Young Talents of Science and Technology in Universities of Inner Mongolia Autonomous Region(NJYT23018)
Innovative Project of Young “Grasslands Talents”
Fundamental Research Funds for the Inner Mongolia Normal University(2022JBBJ013)
Fundamental Research Funds for the Inner Mongolia Normal University(2022JBTD011)
The increasing frequency of recent droughts has an adverse effect on the ecosystem of the Mongolian Plateau. The growth condition of NPP is considered an indicator of the ecological function. Therefore, identifying the relationship between NPP and drought can assist in the prevention of drought-associated disasters and the conservation of the ecological environment of the Mongolian Plateau. This study used the Carnegie-Ames-Stanford Approach (CASA) model to simulate the NPP capacity of the Mongolian Plateau between 1982 and 2015, as well as drought indicators (drought probability, vulnerability, and risk) to explore the drought risk of NPP. The findings pointed to an overall increase in NPP with regional variances; however, the NPP rate in Inner Mongolia was considerably higher than that in Mongolia. The standardized precipitation evapotranspiration index (SPEI) showed an overall downward trend, with Inner Mongolia experiencing a substantially lower rate of decline than Mongolia. The areas most likely to experience drought were primarily in the center and north while the areas with the highest drought vulnerability were primarily in the northeast, center, and southeast. Mongolia showed a higher probability of drought compared to Inner Mongolia. Drought-prone regions of the Mongolian Plateau increased during the 21st century while drought-vulnerable areas increased and shifted from north to south. Alpine grasslands and coniferous forests were least vulnerable to drought, while other vegetation types experienced temporal variation. In the 21st century, the primary determinants of drought risk shifted from precipitation and the normalized difference vegetation index (NDVI) to temperature and relative humidity.
Key words: drought probability; drought vulnerability; drought risk; NPP; Mongolian Plateau
REN Jinyuan , GUO Xiaomeng , TONG Siqin , BAO Yuhai , BAO Gang , HUANG Xiaojun . Risk posed to vegetation net primary productivity by drought on the Mongolian Plateau[J]. Journal of Geographical Sciences, 2023 , 33(11) : 2175 -2192 . DOI: 10.1007/s11442-023-2171-1
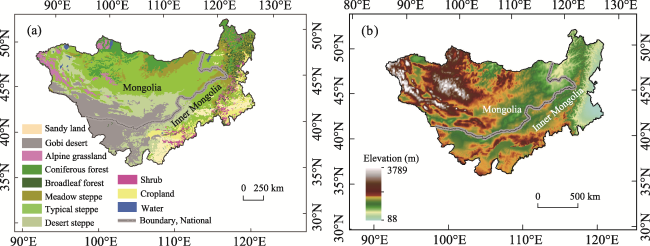
Figure 1 Map of different vegetation types, geographical location (a), and elevation (b) of the Mongolian Plateau |
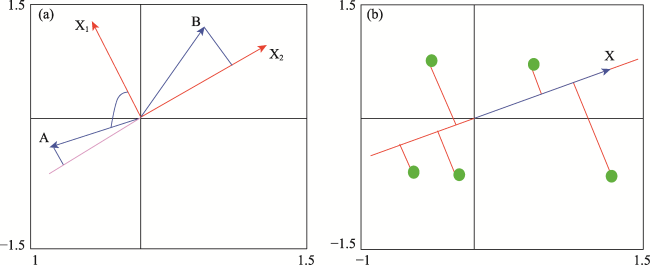
Figure 2 Example diagrams for redundancy analysis, different environmental variables (a), and the sample symbols (b) |
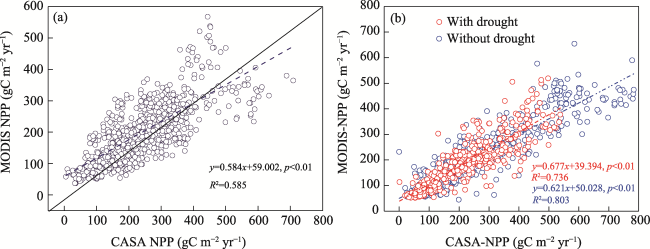
Figure 3 Comparison of the CASA model pixel-scale simulations for average annual scale (a), drought and without drought conditions (b). (Red: with drought, SPEI-12 ≤ −1.0; Blue: without drought, SPEI-12 > −1.0) |
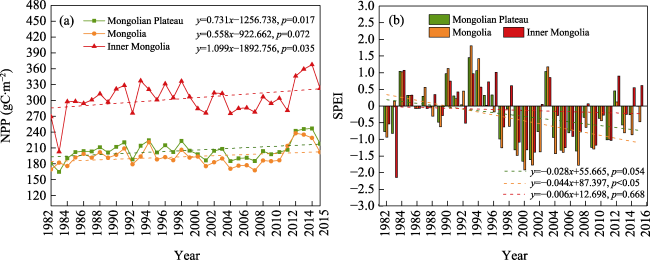
Figure 4 Temporal variations in NPP (a) and SPEI (b) for the Mongolian Plateau |
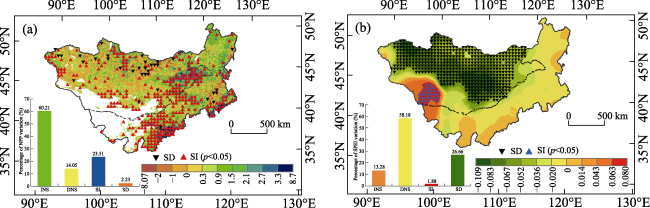
Figure 5 Spatial variations in NPP (a) and SPEI (b) (SI: significant increase; SD: significant decrease; INS: non-significant increase; DNS: non-significant decrease) |
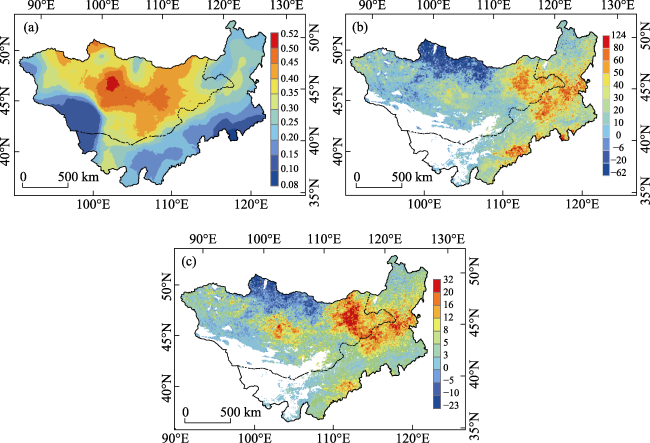
Figure 6 Drought probability (a), vulnerability (b), and risk (c) of NPP on the Mongolian Plateau during 1982- 2015 |
Table 1 Variations of net primary productivity (NPP) and each drought index in different vegetation zones across various periods (I: 1982-2015; II: 1982-1999; III: 2000-2015) |
| Vegetation type | NPP | Drought probability | Drought vulnerability of NPP | Drought risk of NPP | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ⅰ | Ⅱ | Ⅲ | Ⅰ | Ⅱ | Ⅲ | Ⅰ | Ⅱ | Ⅲ | Ⅰ | Ⅱ | Ⅲ | |
| Coniferous forest | 381.12 | 380.06 | 382.32 | 0.35 | 0.19 | 0.54 | -0.02 | 8.97 | -11.41 | -0.41 | 1.62 | -6.86 |
| Broadleaf forest | 638.74 | 642.10 | 634.96 | 0.26 | 0.21 | 0.31 | 38.34 | 72.91 | 11.90 | 9.60 | 16.31 | 2.76 |
| Meadow steppe | 374.12 | 376.94 | 370.96 | 0.32 | 0.20 | 0.47 | 26.22 | 27.29 | 9.83 | 8.07 | 5.76 | 3.83 |
| Typical steppe | 221.44 | 221.14 | 221.78 | 0.35 | 0.18 | 0.53 | 22.94 | 13.18 | 12.89 | 7.47 | 2.35 | 5.20 |
| Desert steppe | 84.55 | 81.19 | 88.32 | 0.31 | 0.18 | 0.46 | 12.44 | 14.32 | 10.94 | 3.84 | 2.56 | 4.58 |
| Shrub | 244.32 | 243.10 | 245.68 | 0.20 | 0.17 | 0.24 | 30.10 | 21.52 | 27.03 | 6.22 | 4.11 | 6.11 |
| Sandy land | 213.66 | 207.49 | 220.61 | 0.19 | 0.15 | 0.22 | 28.27 | 10.24 | 26.91 | 5.31 | 1.66 | 6.01 |
| Cropland | 268.67 | 263.07 | 274.97 | 0.19 | 0.16 | 0.21 | 37.63 | 16.85 | 37.05 | 7.15 | 3.28 | 7.57 |
| Alpine grassland | 203.94 | 202.14 | 205.96 | 0.29 | 0.20 | 0.40 | 9.31 | 6.46 | 3.59 | 2.02 | 0.84 | -0.06 |
| Mongolian Plateau | 239.04 | 237.84 | 240.39 | 0.30 | 0.18 | 0.43 | 20.34 | 16.50 | 12.04 | 5.80 | 3.15 | 3.78 |
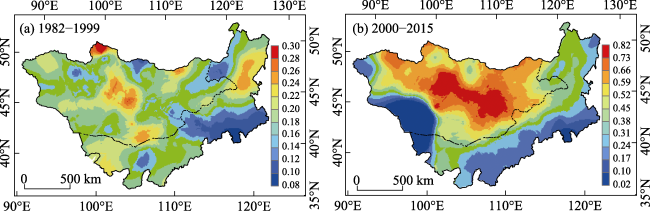
Figure 7 Drought probability on the Mongolian Plateau during 1982-1999 (a) and 2000-2015 (b) |
Figure 8 The drought vulnerability of NPP on the Mongolian Plateau during 1982-1999 (a) and 2000-2015 (b) |
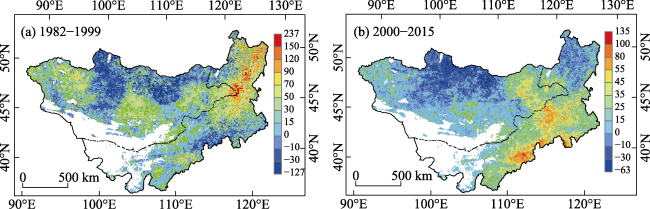
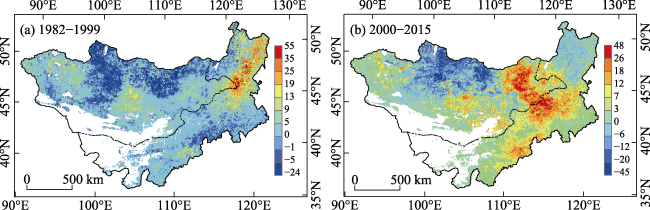
Figure 9 The drought risk of NPP on the Mongolian Plateau during 1982-1999 (a) and 2000-2015 (b) |
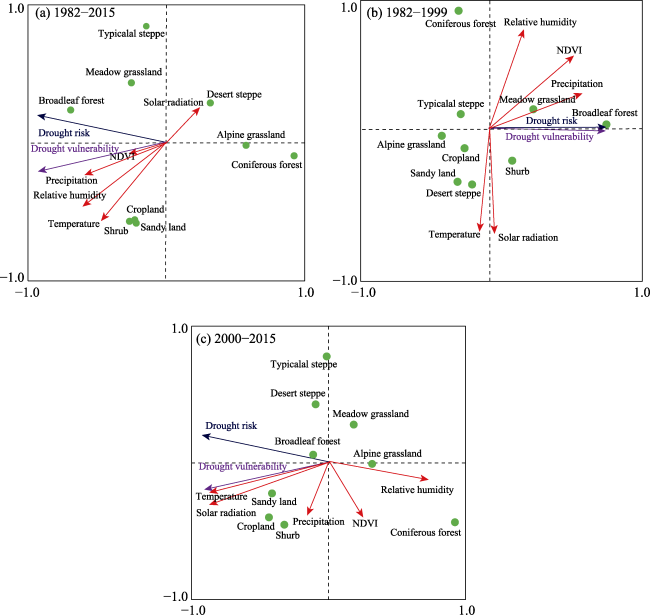
Figure 10 Redundancy analysis of the drought risk and vulnerability of net primary productivity (NPP) during 1982-2015 (a), 1982-1999 (b), and 2000-2015 (c) |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}