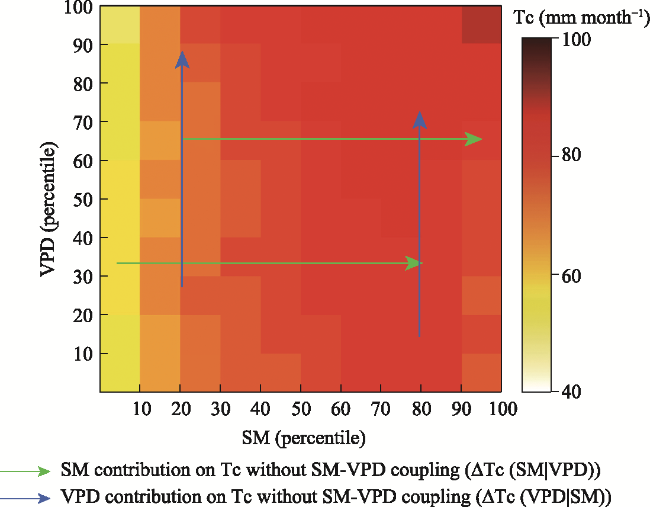
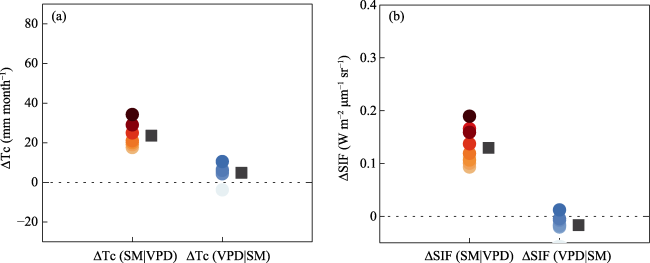
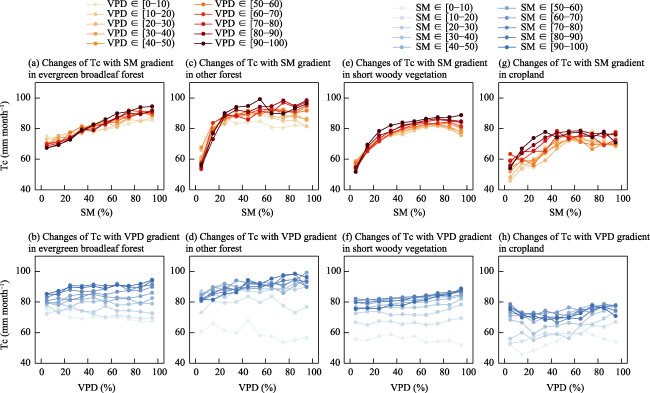
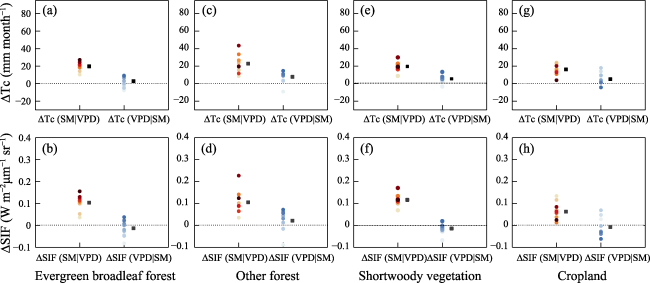
Understanding the driving role of VPD and SM on transpiration changes in the context of climate warming remains to be improved mainly because SM and VPD are coupled through land-atmosphere interactions, restricting the quantification of the independent effects of SM and VPD on Tc. The “binning method” decouples the strong coupling between SM and VPD at regional and global scales (Liu
et al.,
2020; Cheng
et al.,
2022; Yu
et al.,
2022). In contrast, traditional methods such as correlation analysis and multiple linear regression do not sufficiently decouple the covariance between SM and VPD (Liu
et al.,
2020). The response of Tc to the SM and VPD may be nonlinear (Sage and Kubien,
2007; Green
et al.,
2019), and this method can capture this nonlinearity (Dang
et al.,
2022; Fu
et al.,
2022a). The SM-VPD correlation in this study showed a significant decrease after using this method (
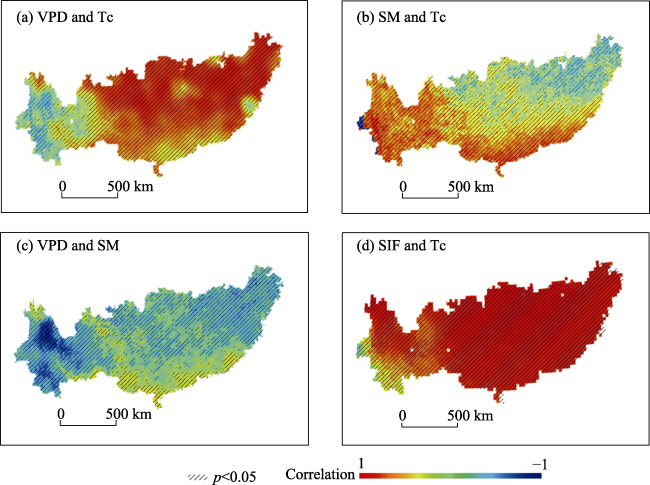
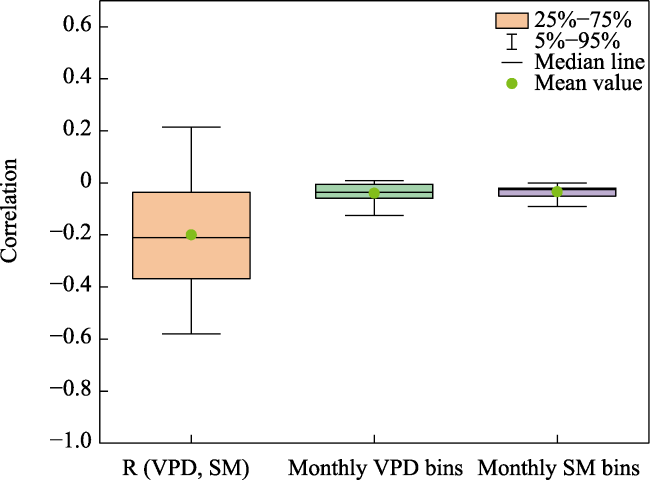
Figure 4), which provided a basis for exploring the independent effects of SM and VPD on Tc. In this study, we analyzed the independent effects of SM and VPD on Tc in subtropical China and found that the independent effects of SM on Tc were greater than those of VPD. This supports the results of Liu
et al. (
2020) and Yu
et al. (
2022) and emphasizes the importance of SM on Tc. Tc is more susceptible to SM in subtropical China, where droughts are frequent and SM highly fluctuates annually (Yuan
et al.,
2016). However, recent studies on the independent effects of SM and VPD on Tc in subtropical China have shown that VPD and SM limit Tc by 90.8% and 9.2% (Song
et al.,
2020), respectively; the reason for this difference might be the different study timescales. Because solar radiation, VPD, and temperature affect transpiration (Granier
et al.,
2000; Bai
et al.,
2015), we excluded pixels with temperatures less than 15℃, VPD less than 5 hPa, and photosynthetic photon flux densities less than 500 µmol m
-2 s
-1 from the study area to control the interference of other factors in the study (Liu
et al.,
2020). Therefore, this study focused on the relative contributions of changes in VPD and SM to Tc during the growing season (mainly summer). However, Song considered the ranges of VPD and SM seasonal variations in Tc throughout the year. Considering the similarity in the seasonal patterns of VPD and Tc in the subtropical regions of China, the correlation between VPD and Tc is higher on the intra-annual scale which shows the dominant role of VPD on Tc to some extent. Recent studies have demonstrated the existence of critical thresholds for plant water stress in terrestrial ecosystems (Fu
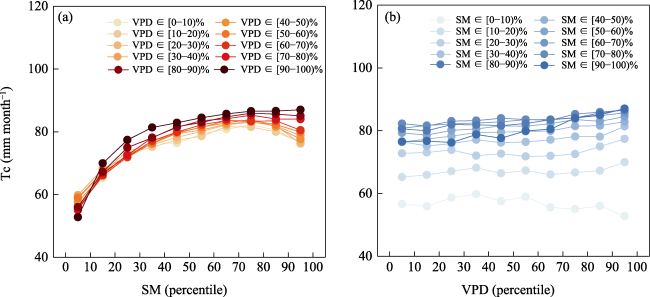
et al.,
2022b,
2022c). The results of this study showed that the response of Tc to SM in the study area was below the 35% threshold, where its increase contributed significantly to Tc. However, above the 35% threshold, changes in SM had a weaker effect on Tc. This indicates that there is a threshold effect on the response of vegetation to SM. Notably, although the threshold response of the VPD was not obvious, a high VPD promoted Tc when the SM was greater than the 10% threshold. Conversely, when the SM was less than the 10% threshold, a high VPD restricted the increase in Tc, indicating that the response of Tc to VPD was regulated by the SM (Liu and Biondi,
2020; Zhang
et al.,
2021). The high VPD in other forests was not always limited to Tc (
Figure 7d), which might be due to the deep, vigorous root systems of forests that could still utilize deep soil water under atmospheric drought (McDowell
et al.,
2008; Zhang
et al.,
2020). In contrast, croplands have a greater tendency to close their stomata to reduce water consumption under water stress, because of their shallow root systems (Chen
et al.,
2021) (
Figure 7h). High VPD and SM values contributed to Tc in forests under the same SM or VPD conditions (
Figure 8a). However, croplands showed the opposite trend (
Figure 8g), which may be related to environmental conditions, soil properties, and root systems across forests, short woody vegetation, and croplands (Chen
et al.,
2021). In future studies, it should distinguish the sensitivity of different vegetation types to SM and VPD in ecosystem water cycle simulations. In addition, numerous recent studies have shown that SIF is closely related to Tc and can be used to represent changes in Tc (Pagán
et al.,
2019; Feng
et al.,
2021; Shan
et al.,
2021). Figures 3d and 8 show that using SIF is feasible, verifying the feasibility of our analysis and the reasonableness of the results, respectively.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}