
Journal of Geographical Sciences >
Human sedentism and use of animal resources on the prehistoric Tibetan Plateau
|
Wang Qing (1998-), PhD Candidate, specialized in environmental archaeology. E-mail: wangqing19@mails.ucas.ac.cn |
Received date: 2023-04-05
Accepted date: 2023-06-20
Online published: 2023-10-08
Supported by
National Natural Science Foundation of China(41930323)
The Second Tibetan Plateau Scientific Expedition and Research Program(2019QZKK0601)
The transition of human societies from high mobility to sedentary lifestyles had a profound impact on subsistence, technology, and the origin of civilization. Sedentism was influenced by various factors such as climate change, population growth, resource pressure, and technological innovation. The Tibetan Plateau, due to its alpine and hypoxic conditions, is an ideal region to study human adaptation to extreme environments. However, the prehistoric process of sedentism on the Tibetan Plateau is unclear and the chronological sequence and driving mechanism of sedentism on the Tibetan Plateau are still controversial. Previous studies have focused on the diffusion of agriculture from low to high elevation areas, with little attention given to the role of animal resources in sedentism. Seasonality analysis using animal remains is crucial in determining whether a site was occupied year-round. To establish the seasonal calendar of animal resource utilization, it is recommended to create a database of skeletal morphology, whole genome, and proteome of contemporary Tibetan Plateau fauna to aid in the identification of animal remains from archaeological sites. Thus, intricate web of human-animal-environment relationship and the role of animal resources in human sedentism on the Tibetan Plateau can then be evaluated.
WANG Qing , ZHANG Ying , CHEN Shungang , GAO Yu , YANG Jishuai , RAN Jingkun , GU Zhengquan , YANG Xiaoyan . Human sedentism and use of animal resources on the prehistoric Tibetan Plateau[J]. Journal of Geographical Sciences, 2023 , 33(9) : 1851 -1876 . DOI: 10.1007/s11442-023-2156-0
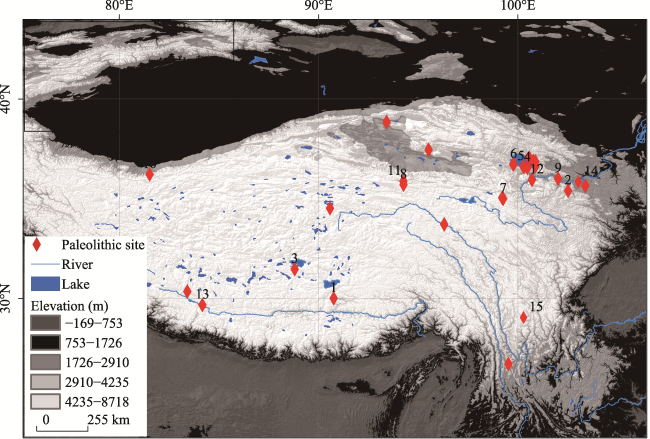
Figure 1 The distribution of Paleolithic sites on the Tibetan Plateau (1. Chusang (226-169 ka BP, 4369 m a.s.l.; Zhang et al., 2021); 2. Baishiya Karst Cave (190-5 ka BP, 3200 m a.s.l.; Chen et al., 2019; Zhang et al., 2020); 3. Nwya Devu (Zhang et al., 2018); 4. 151 (Wang et al., 2020); 5. Jiangxigou#1; 6. Heimahe#1 (13.1 ka BP, 3200 m a.s.l.; Brantingham, 2006); 7. Xiadawu (~11 ka BP, 3988 m a.s.l.; Hou et al., 2016); 8. Xidatan#2 (Brantingham et al., 2013); 9. Shalongka (8.3-8 ka BP, 2046 m a.s.l.; Dong et al., 2013); 10. Yangchang (7.6-7 ka BP, 2440 m a.s.l.; Han et al., 2014); 11. Yeniugou (7.5 ka BP, 3800 m a.s.l.; Tang et al., 2013); 12. Layihai (~7.6 ka BP, 3125 m a.s.l.; Gai and Wang, 1983); 13. Zhongba 10-1 (6.6-2.6 ka BP, 4570 m a.s.l.; Hudson et al., 2014); 14. Jiangjunfu 01 (120-90 ka BP, 2673 m a.s.l.; Cheng et al., 2021); 15. Piluo (~130 ka BP, 3750 m a.s.l.; Zheng et al., 2022) |
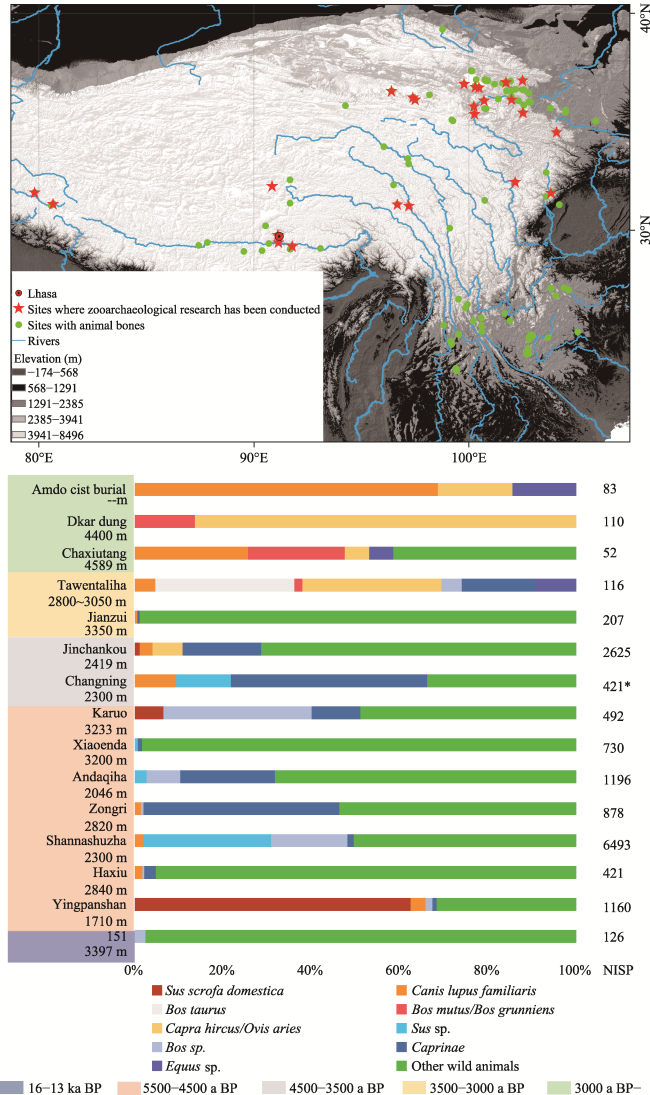
Figure 2 The archaeological sites with animal remains unearthed on the Tibetan Plateau and their proportions of animals (annotation: left column gives the names and elevations of the sites in chronological order, horizontal bars show the percentage of animal types recorded for each site, and the right column gives the Number of Identified Specimens (NISP) at the sites; * is MNI the Minimum Number of Individuals at Changning site.)) |
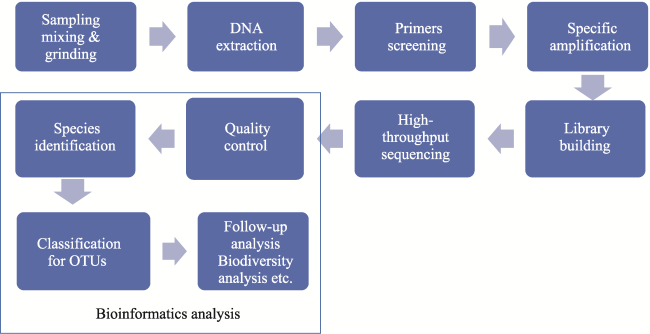
Figure 3 Experimental workflow of bulk-bone metabarcoding (OTU = operational taxonomic unit) |
Table 1 Primers for specific amplification in bulk-bone metabarcoding |
| Name | Primer (5°-3°) | Target taxa | Target gene | Reference | Amplicon length (BP) | Annealing temper- ature (℃) | |
|---|---|---|---|---|---|---|---|
| 12SAH | Forward | CTGGGATTAGATACCCCACTAT | Bird | 12S rRNA | Cooper, 1994 | 229-235 | 57 |
| Reverse | CCTTGACCTGTCTTGTTAGC | Bird | 12S rRNA | Cooper, 1994 | 229-235 | 57 | |
| Mam16S | Forward | CGGTTGGGGTGACCTCGGA | Mammal | 16S rRNA | Taylor, 1996 | 89-115 | 57 |
| Reverse | GCTGTTATCCCTAGGGTAACT | Mammal | 16S rRNA | Taylor, 1996 | 89-115 | 57 | |
| 12S-V5 | Forward | ACTGGGATTAGATACCCC | Fish | 12S rRNA | Riaz et al., 2011 | 106 | 58 |
| Reverse | TAGAACAGGCTCCTCTAG | Fish | 12S rRNA | Riaz et al., 2011 | 106 | 58 |
Table 2 Completeness of mitochondrial DNA data of living animals on the Tibetan Plateau |
| Taxa | Number of recorded species | Number of counted species | MtDNA database | Notes | ||
|---|---|---|---|---|---|---|
| Complete | Partial | None | ||||
| Mammal | ca. 190 | 149 | 119 | 15 | 13 | The animal species that lack mtDNA data are primarily Vespertilionidae, Muridae, Columbidae, and Caprimulgidae, and only Moschus fuscus in Artiodactyla has no complete mtDNA data |
| Fish | 159 | 152 | 112 | 20 | 20 | Species without data are distributed in all families, but other species of the same genus always have data |
| Bird | 817 | 234 | 152 | 71 | 11 | Most of the data are not available for Rallidae, Scolopacidae, Scolopacidae, and Caprimulgidae among others. |
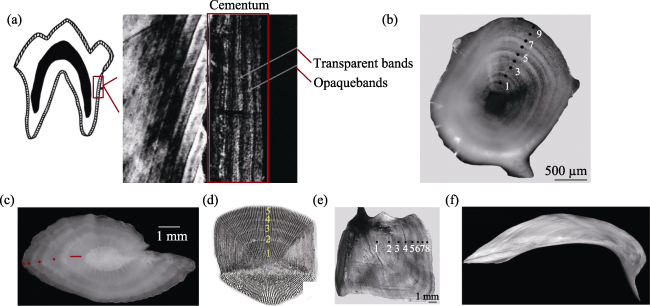
Figure 4 Animal bone materials used for seasonality analysis (a. Position of cementum and microstructure of cementum under polarizing microscope (Fišáková, 2013); b. fish vertebra (Ma et al., 2017); c. otolith (Van Neer et al., 1999); d. fish scale; e. opercular (Ma et al., 2017); f. cleithrum (Faust et al., 2013)) |
Table 3 Timing of the winter-summer boundary of dominant fish in the middle reaches of Yarlung Zangbo |
| Species | Number of ring cycles per year | Month | Analyzed material | Reference |
|---|---|---|---|---|
| Ptychobarbus dipogon | 1 | 3-5 | Otolith | Li et al., 2009 |
| 1 | 3-6 | Otolith | Yang, 2015 | |
| Oxygymnocypris stewartii | 1 | 3-6 | Otolith and vertebra | Huo, 2014 |
| 3-5 | ||||
| Schizopygopsis younghusbandi | 1 | 3-5 | Otolith | Duan, 2015 |
| Glyptosternum maculatum | 1 | 4-6 | Vertebra | Ding et al., 2008 |
| Schizothorax o’connori | 1 | 3-5 | Otolith | Ma et al., 2011 |
| 3-6 | Vertebra | |||
| Schizothorax waltoni | 1 | 3-5 | Otolith and vertebra | Hao, 2005; |
| Zhou, 2014 | ||||
| Schizothorax macropogon | 1 | 3-6 | Otolith | Liu, 2016 |
| Gymnocypris waddellii | 1 | - | Anal scale and dorsal fin ray | Yang et al., 2011 |
| [1] |
Administration Commission of Cultural Relics of Tibet Autonomous Region ACCRTAR, 1985. Karuo Site in Chamdo Region. Beijing: Cultural Relics Publishing House. (in Chinese)
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
Chengdu Municipal Institute of Cultural Relics and Archaeology CMICRA, Administration of Cultural Relics of Aba Tibetan and Qiang Autonomous Prefecture, Qiang Museum of Maoxian County, 2018. The Yingpanshan Neolithic Site in Maoxian Couty. Beijing: Cultural Relics Publishing House. (in Chinese)
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
Heritage Survey Team of Administration Commission of Cultural Relics of Tibet Autonomous Region HSTACCRTAR, 1990. Trial excavation report of Xiaoenda Neolithic site in Tibet Autonomous region. Archaeology and Cultural Relics, (1): 28-43. (in Chinese)
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
Institute of Archaeology, Chinese Academy of Social Sciences IACASS, 1999. Qugong Site in Lhasa. Beijing: Encyclopedia of China Publishing House. (in Chinese)
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|
| [114] |
|
| [115] |
|
| [116] |
|
| [117] |
|
| [118] |
|
| [119] |
|
| [120] |
|
| [121] |
|
| [122] |
|
| [123] |
|
| [124] |
|
| [125] |
|
| [126] |
|
| [127] |
|
| [128] |
|
| [129] |
Sichuan Provincial Cultural Relics and Archaeology Research Institute, Administration of Cultural Relics of Aba Tibetan and Qiang Autonomous Prefecture, Administration of Cultural Relics of Jinchuan County Archaeology, 2021. 2011 Excavation of Liujiazhai Site in Jinchuan County, Sichuan. Archaeology, (3): 16. (in Chinese)
|
| [130] |
|
| [131] |
|
| [132] |
|
| [133] |
|
| [134] |
|
| [135] |
|
| [136] |
|
| [137] |
Tibet Institute for Conservation and Research of Cultural Relics TICRCR, Shaanxi Provincial Institute of Archaeology, Zada County Bureau of Culture, 2022. Brief report of 2017 annual archaeological excavation in Gebuseru Cemetery, Alizada County, Tibet. In: Li L H. Archaeological Research on Tibetan Cultural Relics (Series 4). Beijing: Science Press: 1-27.
|
| [138] |
Tibetan Zoology Compilation Leading Group TZCLG, 2005. The Chorography of Tibet Autonomous Region:Fauna. Beijing: China Tibetology Publishing House, 588. (in Chinese)
|
| [139] |
|
| [140] |
|
| [141] |
|
| [142] |
|
| [143] |
|
| [144] |
|
| [145] |
|
| [146] |
|
| [147] |
|
| [148] |
|
| [149] |
|
| [150] |
|
| [151] |
|
| [152] |
|
| [153] |
|
| [154] |
|
| [155] |
|
| [156] |
|
| [157] |
|
| [158] |
|
| [159] |
|
| [160] |
|
| [161] |
|
| [162] |
|
| [163] |
|
| [164] |
|
| [165] |
|
| [166] |
|
| [167] |
|
| [168] |
|
| [169] |
|
| [170] |
|
| [171] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}