
Journal of Geographical Sciences >
Considering time-lag effects can improve the accuracy of NPP simulation using a light use efficiency model
|
Li Chuanhua, Associate Professor, specialized in ecological remote sensing. E-mail: lch_nwnu@126.com |
Received date: 2022-04-24
Accepted date: 2022-09-26
Online published: 2023-05-11
Supported by
National Natural Science Foundation of China(42161058)
The State Key Laboratory of Cryospheric Science(SKLCS-ZZ-2022)
The West Light Foundation of the Chinese Academy of Sciences
Most terrestrial models synchronously calculate net primary productivity (NPP) using the input climate variable, without the consideration of time-lag effects, which may increase the uncertainty of NPP simulation. Based on Normalized Difference Vegetation Index (NDVI) and climate data, we used the time lag cross-correlation method to investigate the time-lag effects of temperature, precipitation, and solar radiation in different seasons on NDVI values. Then, we selected the Carnegie-Ames-Stanford approach (CASA) model to estimate the NPP of China from 2002 to 2017. The results showed that the response of vegetation growth to climate factors had an obvious lag effect, with the longest time lag in solar radiation and the shortest time lag in temperature. The time lag of vegetation to the climate variable showed great tempo-spatial heterogeneities among vegetation types, climate types, and vegetation growth periods. Based on the validation using eddy covariance data, the results showed that the simulation accuracy of the CASA model considering the time-lag effects was effectively improved. By considering the time-lag effects, the average total amount of NPP modeled by CASA during 2001-2017 in China was 3.977 PgC a-1, which is 11.37% higher than that of the original model. This study highlights the importance of considering the time lag for the simulation of vegetation growth, and provides a useful tool for the improvement of the vegetation productivity model.
LI Chuanhua , LIU Yunfan , ZHU Tongbin , ZHOU Min , DOU Tianbao , LIU Lihui , WU Xiaodong . Considering time-lag effects can improve the accuracy of NPP simulation using a light use efficiency model[J]. Journal of Geographical Sciences, 2023 , 33(5) : 961 -979 . DOI: 10.1007/s11442-023-2115-9
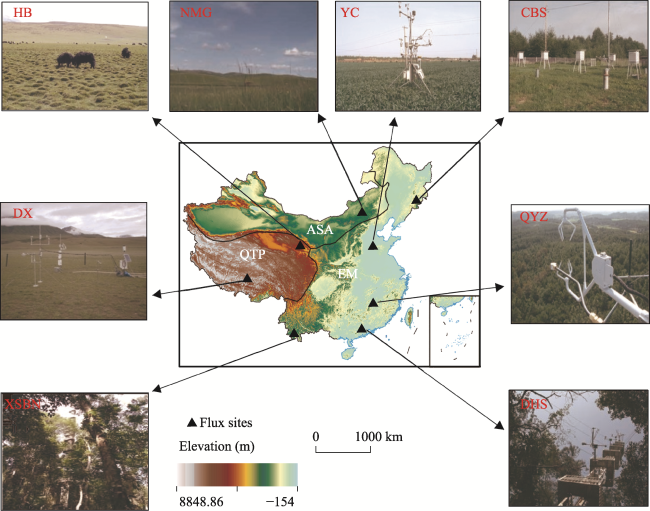
Figure 1 Overview of the three climatic regions of China |
Table 1 Geographic coordinates, NPP/GPP values (φ), and vegetation types for the eddy covariance sites |
| Sites | Longitude | Latitude | φ | Types |
|---|---|---|---|---|
| CBS | 128°06′E | 42°24′N | 0.5488 | Forest |
| QYZ | 115°03′E | 26°44′N | 0.4125 | Forest |
| DHS | 112°30′E | 23°09′N | 0.4125 | Forest |
| XSBN | 101°16′E | 21°54′N | 0.4125 | Forest |
| NMG | 116°18′E | 44°08′N | 0.4000 | Grass |
| HB | 101°20′E | 37°40′N | 0.5523 | Grass |
| DX | 91°03′E | 30°29′N | 0.5523 | Grass |
| YC | 116°38′E | 36°58′N | 0.5399 | Crops |
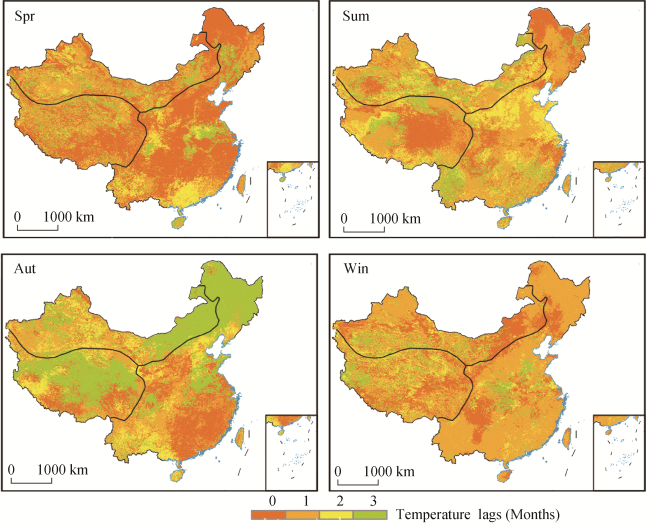
Figure 2 The average time lag of the vegetation response to temperature in different seasons during 2001-2017 |
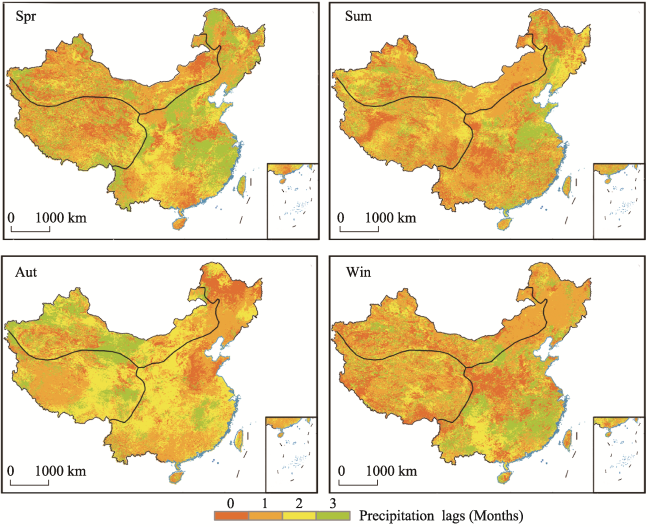
Figure 3 The average time lag of the vegetation response to precipitation in different seasons during 2001-2017 |
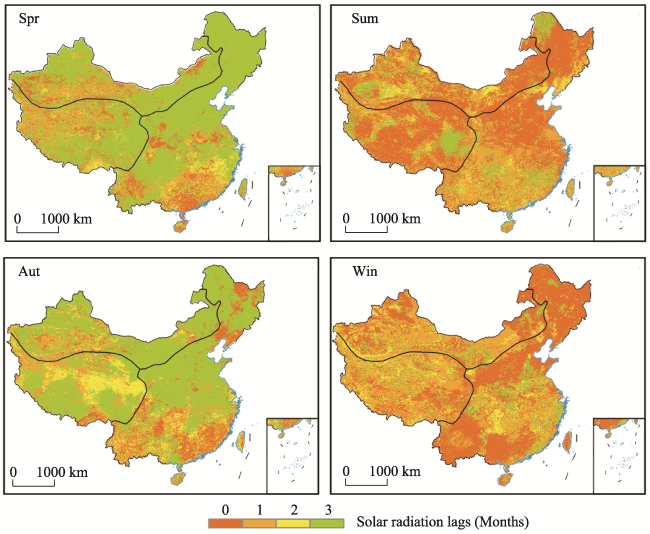
Figure 4 The averaged lag time of the vegetation response to solar radiation in different seasons during 2001-2017 |
Table 2 Time-lag period of climatic factors in the different climate zones and vegetation types |
| Temperature | Precipitation | Solar radiation | |
|---|---|---|---|
| China | 1.24 | 1.49 | 1.58 |
| EM | 1.11 | 1.53 | 1.50 |
| QTP | 1.30 | 1.49 | 1.69 |
| ASA | 1.40 | 1.43 | 1.60 |
| Farmland | 1.18 | 1.50 | 1.52 |
| Forest | 1.06 | 1.54 | 1.51 |
| Grassland | 1.26 | 1.42 | 1.67 |
| Desert | 1.45 | 1.54 | 1.59 |
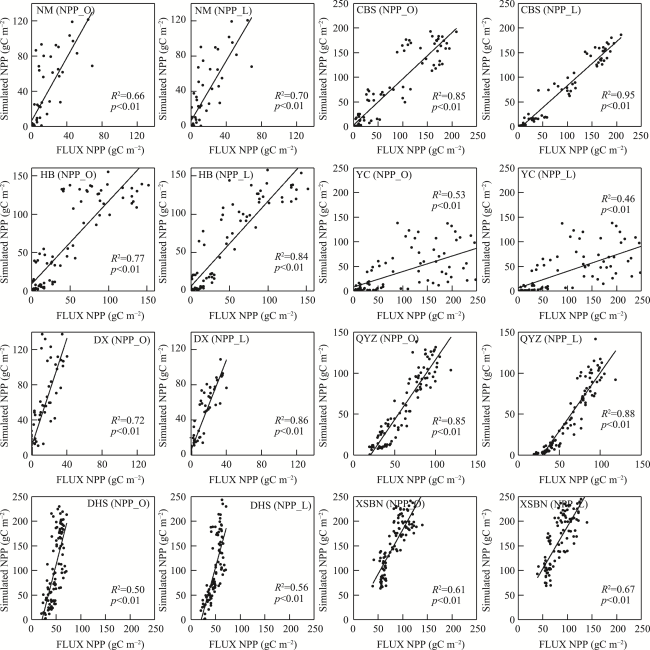
Figure 5 Comparison of the eddy flux site data and the simulated NPP during 2003-2010 |
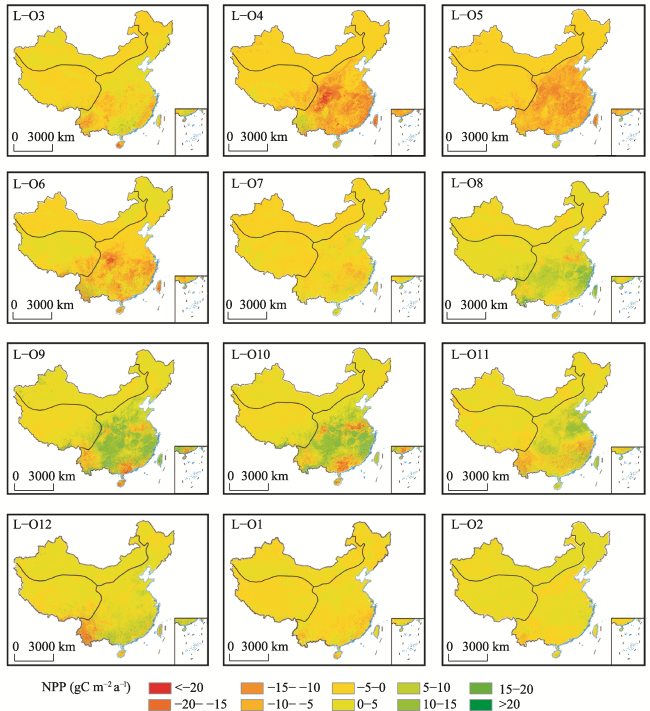
Figure 6 Average monthly difference in NPP_L minus NPP_O from January to December 2001-2017 |
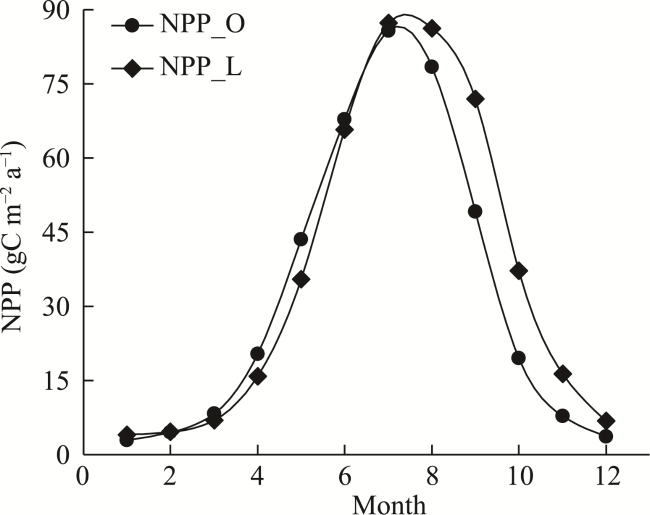
Figure 7 The average monthly changes in NPP from NPP_O and NPP_L during 2001-2017 |
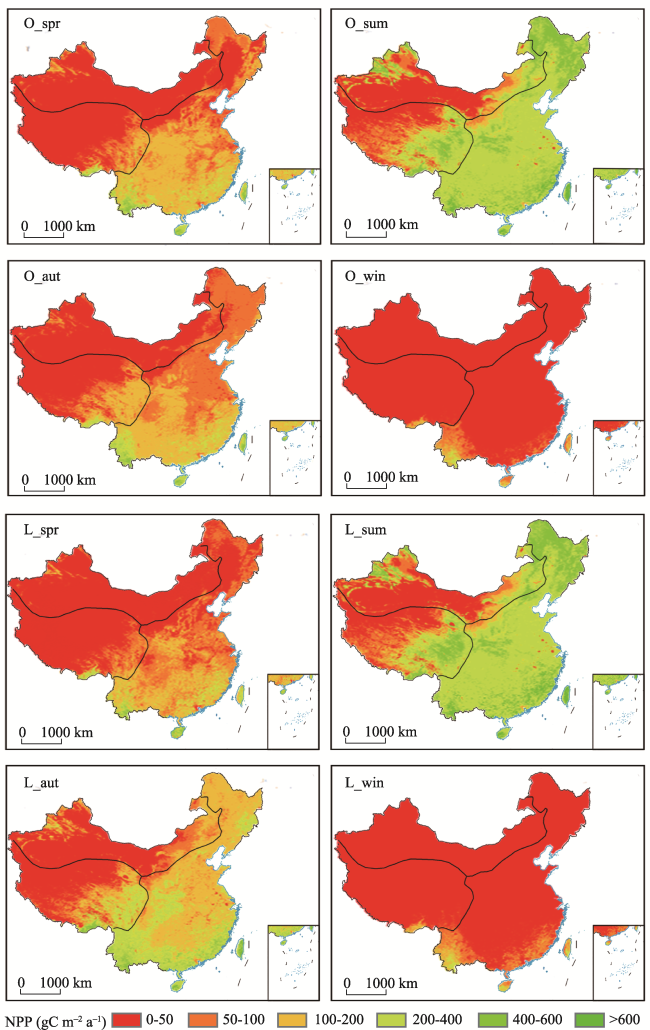
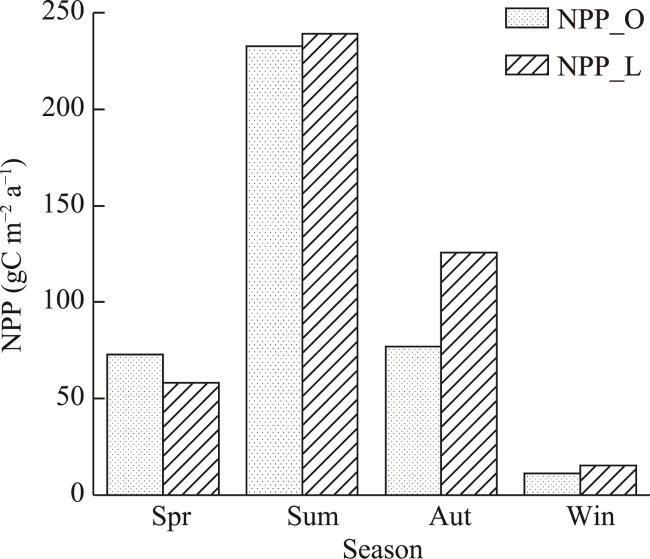
Figure 8 The average seasonal NPP values from NPP_O and NPP_L during 2001-2017 (O indicates the simulated NPP using the model without consideration of time-lag effects, and L indicates the simulated NPP values using the model that considered time-lag effects; spr, sum, aut, and win are abbreviations for the respective seasons.) |
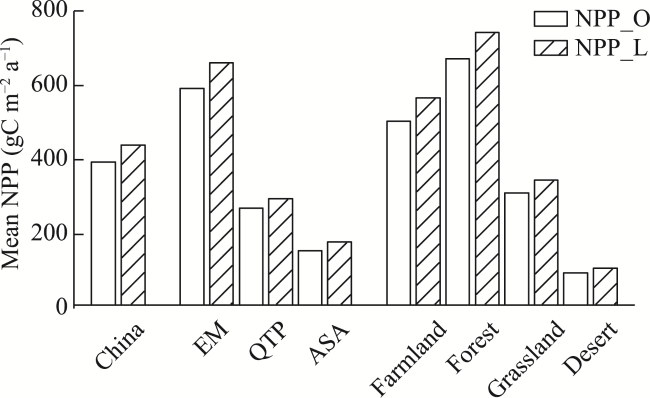
Figure 9 The average seasonal NPP values from NPP_O and NPP_L for China’s terrestrial ecosystems during 2001-2017 (O indicates the simulated NPP using the model without consideration of time-lag effects, and L indicates the simulated NPP values using the model that considered time-lag effects; spr, sum, aut, and win are abbreviations for the respective seasons.) |
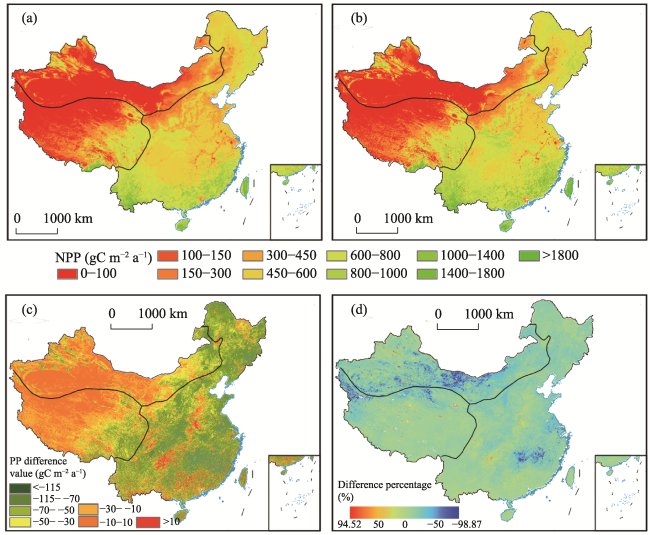
Figure 10 Comparison between the average annual NPP values from the NPP_O and NPP_L models during 2001-2017 (Spatial distribution of the NPP values from NPP_O (a) and NPP_L (b); the absolute (c) and relative (d) differences between the NPP values from the NPP_O minus NPP_L models) |
Figure 11 Comparison of the annual NPP values from NPP_O and NPP_L of China’s terrestrial ecosystems during 2001-2017 (EM indicates the eastern monsoon region, ASA indicates the northwest arid and semi-arid region, and QTP indicates the Qinghai-Tibet Plateau.) |
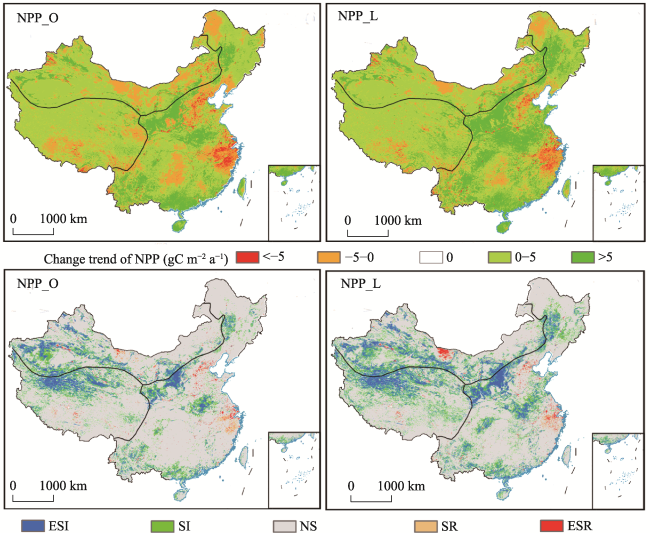
Figure 12 Changing trends and significant test of the annual NPP_O and NPP_L simulated NPP values from 2001 to 2017 (ESI and SI indicate a significant increasing trend at p < 0.01 and p < 0.05; ESR and ER indicate a significant decreasing trend at p < 0.01 and p < 0.05.) |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}