
Journal of Geographical Sciences >
Water sources for typical desert vegetation in the Ebinur Lake basin
|
Hao Shuai (1982-), specialized in ecohydrological process in arid zone. E-mail: haoshuai1869@163.com |
Received date: 2022-01-23
Accepted date: 2022-03-03
Online published: 2022-08-25
Supported by
Natural Science Foundation of Xinjiang Uygur Autonomous Region(2021D01A118)
National Natural Science Foundation of China(U1803244)
Open Project of Xinjiang Laboratory of Lake Environment and Resources in Arid Area(XJNUSYS2019B18)
In arid and semi-arid environments, desert vegetation plays an important role in preventing soil erosion by wind and helps maintain the stability of desert and oasis ecosystems. Four types of typical desert vegetation, namely Populus euphratica, Haloxylon ammodendron, Nitraria sibirica, and Halostachs caspica, corresponding to different habitats (i.e., river bank, sand dune, desert, and salt marsh) were chosen as the model vegetation in this research. The δ2H and δ18O for rainwater, soil water, and plant water were applied to identify the water sources and quantify the proportions of different water sources used over the entire plant growth period (from March to October). The results showed that the precipitation δ2H and δ18O in the Ebinur Lake basin varied from -142.5‰ to -0.6‰ and from -20.16‰ to 1.20‰, respectively. The largest δ2H and δ18O values occurred in summer and the smallest in winter. The soil water δ2H and δ18O of the four habitats decreased gradually with increasing depth. The δ2H and δ18O values of water extracted from the stems of the four plants had similar variation trends, that is, the maximum was observed in spring and the minimum in summer. Among the four plants, H. caspica had the highest stable isotopic values in the stem water, followed by N. sibirica, H. ammodendron, and P. euphratica. The water sources and utilization ratios of desert vegetation varied across different growth stages. Throughout the growing period, H. ammodendron mainly used groundwater, whereas the water source proportions used by N. sibirica varied greatly throughout the growing season. In spring, plants mainly relied on surface soil water, with a contribution rate of 80%-94%. However, in summer, the proportion of deep soil water used was 31%-36%; and in autumn, the proportion of middle soil water used was 33%-36%. H. caspica mainly relied on topsoil water in spring and autumn, and the proportion of soil water in the middle layer slightly increased to 20%-36% in summer. P. euphratica mainly used intermediate soil water in spring with a utilization rate of 53%-54%. In summer, groundwater was the main source, with a utilization rate of 72%-88%, and only 2%-5% came from river water, whereas in autumn, the river water utilization rate rose to 11%-21%. The results indicated that there were significant differences in water use sources during the growing period for desert vegetation in arid areas. This research provides a theoretical basis for understanding water use mechanisms, water adaptation strategies, and vegetation restoration and management in arid areas.
HAO Shuai , LI Fadong . Water sources for typical desert vegetation in the Ebinur Lake basin[J]. Journal of Geographical Sciences, 2022 , 32(6) : 1103 -1118 . DOI: 10.1007/s11442-022-1987-4
Table 1 Morphological characteristics of four desert plants in the Ebinur Lake basin |
| Vegetation | Height (cm) | Crown (cm NS×EW) | Plant type | Foot type | Plant community | Community cover (%) | Habitat | Sample site |
|---|---|---|---|---|---|---|---|---|
| H. caspica | 52.3±12.3 | (21.3±9.2)× (12.1±4.6) | Small shrub | Shallow root | H. caspica- K. foliatum | 15 | Salt marsh | Beidi station |
| P. euphratica | 312.2±24.3 | (214.2±45.6)× (245.4±56.4) | Tree | Middle-deep root | P. euphratica- P. australis | 50 | River bank | Bohe station |
| H. ammodendron | 125.3±14.4 | (56.4±10.3)× (64.5±23.5) | Small tree | Middle-deep root | H. ammodendron | 10 | Sand dune | Tuoniao station |
| N. sibirica | 23.2±9.4 | (104.6±18.3)× (96.3±10.5) | Small shrub | Shallow root | T. ramosissima- N. sibirica | 30 | Desert | Tuoniao station |
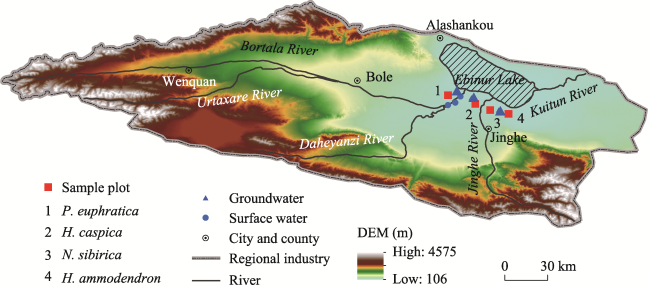
Figure 1 Schematic diagram of sample setting and sampling points in the Ebinur Lake basin |
Table 2 Seasonal variation of precipitation δ2H and δ18O in the Ebinur Lake basin |
| δ(‰) | Spring | Summer | Autumn | Winter | |
|---|---|---|---|---|---|
| δ2H | Average | -96.6 | -45.5 | -103.0 | -129.7 |
| Range | -98.4 to -78.4 | -63.1 to -34.5 | -132.7 to -62.8 | -148.2 to -98.3 | |
| δ18O | Average | -12.33 | -2.50 | -11.31 | -18.63 |
| Range | -12.89 to -9.32 | -5.94 to 1.20 | -17.74 to -5.64 | -20.16 to -14.38 |
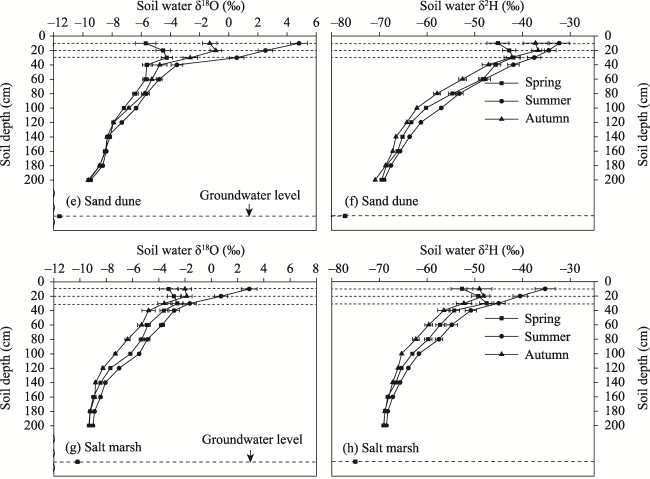
Table 3 Average values of soil water content and soil moisture δ2H and δ18O in different habitats of the Ebinur Lake basin |
| Habitat | 0-60 cm | 60-140 cm | 140-200 cm | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Water content (%) | δ2H (‰) | δ18O (‰) | Water content (%) | δ2H (‰) | δ18O (‰) | Water content (%) | δ2H (‰) | δ18O (‰) | |
| Sand dune | 2.9±1.2 | -41.5±15.2 | -2.48±1.45 | 5.2±1.0 | -60.5±8.1 | -7.19±0.93 | 8.9±1.1 | -66.3±5.2 | -8.50±0.49 |
| Desert | 3.2±1.3 | -51.4±13.7 | -3.40±1.25 | 7.5±1.2 | -67.5±7.3 | -7.88±1.08 | 11.2±1.6 | -73.8±4.7 | -10.21±0.84 |
| River bank | 5.2±1.5 | -55.6±11.4 | -4.31±1.95 | 10.2±1.6 | -73.1±6.7 | -8.82±0.83 | 18.4±2.3 | -79.3±5.7 | -11.20±0.43 |
| Salt marsh | 3.5±1.4 | -48.7±17.1 | -2.37±1.86 | 7.9±2.3 | -63.5±8.2 | -6.90±1.21 | 11.3±3.4 | -68.4±3.8 | -9.02±0.35 |
Note: The data indicate mean value ± standard deviation. |
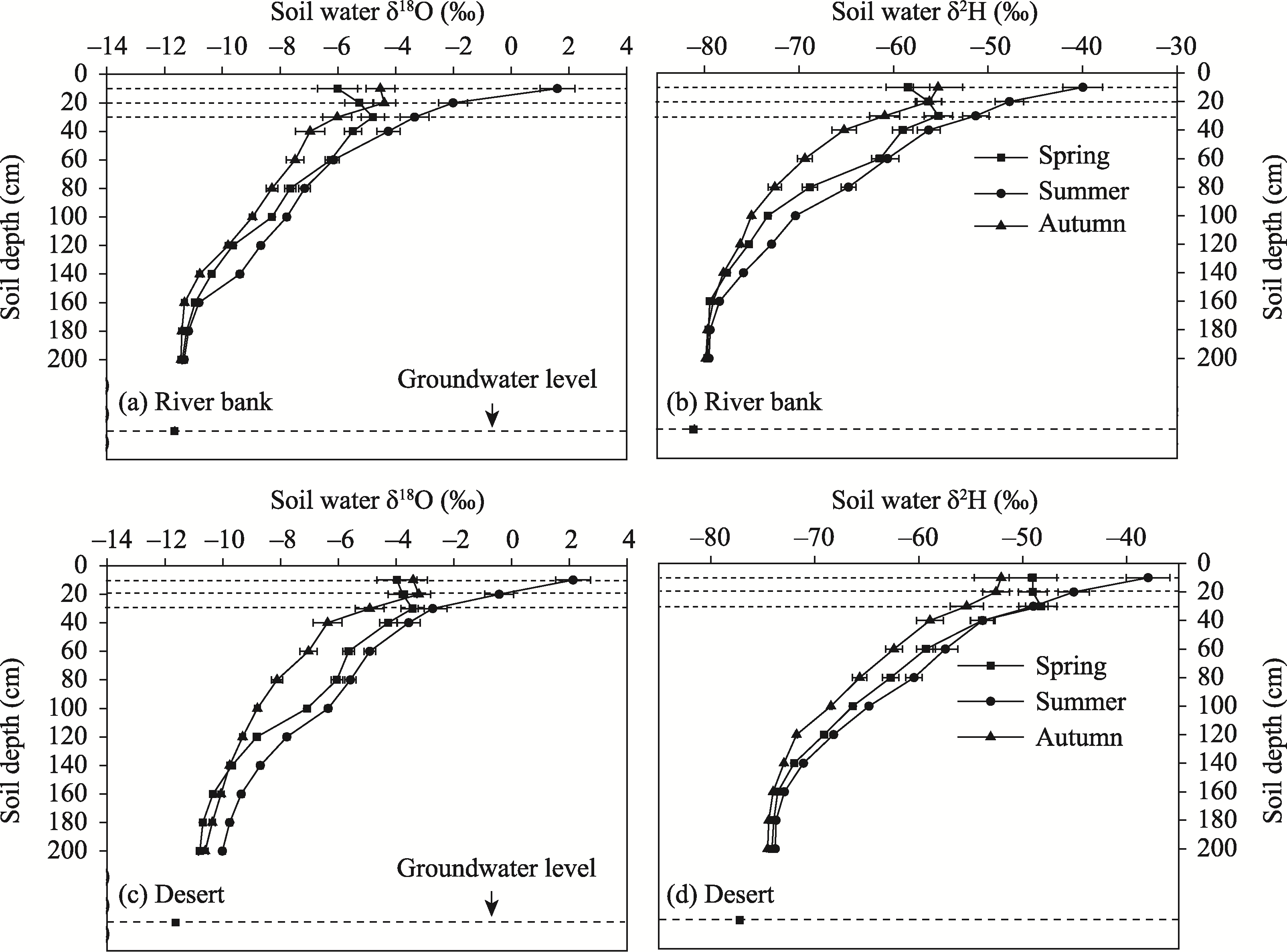
Figure 2 Variations in soil moisture δ2H and δ18O at different depths in different habitats of Ebinur Lake basin |
Table 4 Seasonal variation of δ2H and δ18O values in observation well water of the Ebinur Lake basin |
| Well number | Spring | Summer | Autumn | Well depth (m) | |||
|---|---|---|---|---|---|---|---|
| δ2H (‰) | δ18O (‰) | δ2H (‰) | δ18O (‰) | δ2H (‰) | δ18O (‰) | ||
| 1# | -80.1±2.4 | -12.03±0.92 | -80.5±1.8 | -12.32±0.74 | -80.3±3.1 | -12.25±1.26 | 75 |
| 2# | -74.6±2.5 | -10.44±0.63 | -75.0±2.6 | -10.71±1.12 | -74.8±2.4 | -10.63±0.83 | 80 |
| 3# | -76.8±2.6 | -11.35±1.21 | -77.2±1.6 | -11.63±0.45 | -77.0±2.3 | -11.46±0.71 | 80 |
Note: The data indicate mean value ± standard deviation. |
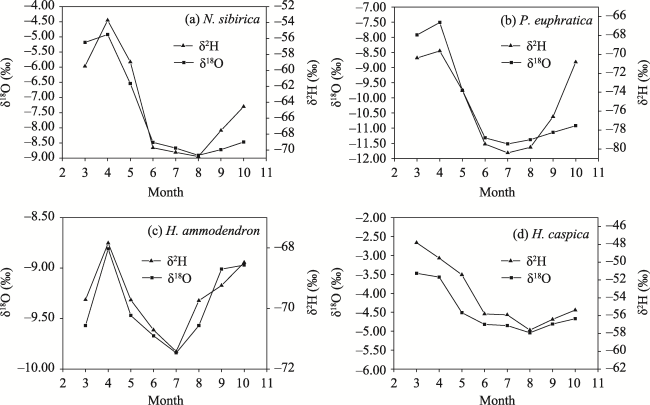
Figure 3 Monthly variation of xylem water δ2H and δ18O in the four plants of the Ebinur Lake basin |
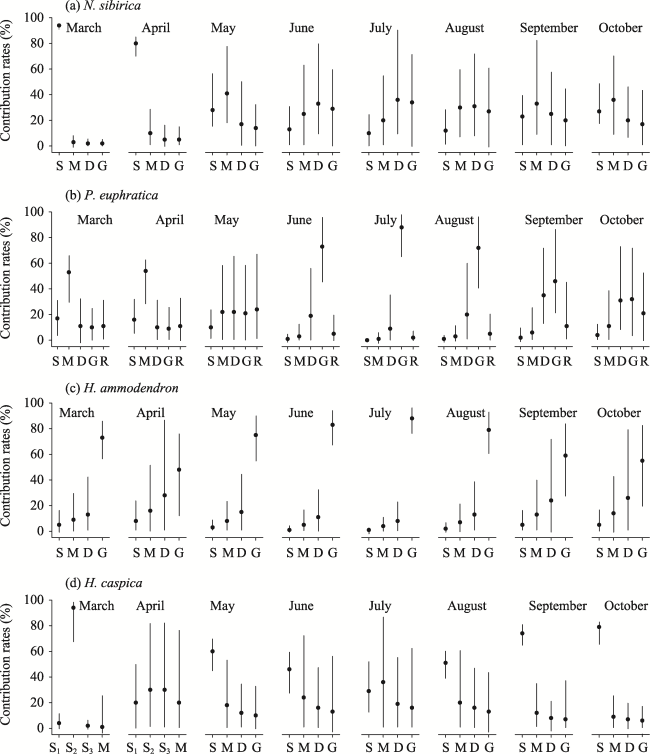
Figure 4 Contribution rates of water consumption from different water sources of the Ebinur Lake basin (S1, S2, S3, S, M, and D represent the soil water in depth of 0-20 cm, 20-40 cm, 40-60 cm, 0-60 cm, 60-140 cm, and 140-200 cm, respectively. G and R represent groundwater and river water. The black dots represent mean values, and the vertical lines represent the range of contribution rates) |
Table 5 Temperature, salinity and pH values of soil profile in different habitats of the Ebinur Lake basin |
| Habitat | 0-20 cm | 20-40 cm | 40-60 cm | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Temperature (℃) | pH | Salinity (g/kg) | Temperature (℃) | pH | Salinity (g/kg) | Temperature (℃) | pH | Salinity (g/kg) | |
| Sand dune | 45.6±3.4 | - | - | 40.3±3.5 | - | - | 32.2±2.9 | - | - |
| Salt marsh | 40.5±3.6 | 9.33±0.34 | 38.3±5.6 | 36.2±2.9 | 8.92±0.25 | 24.5±3.5 | 29.6±2.1 | 8.85±0.14 | 13.5±2.4 |
| Desert | 33.4±4.4 | 8.82±0.21 | 26.5±4.8 | 27.5±3.2 | 8.73±0.15 | 15.6±4.1 | 23.4±2.6 | 8.75±0.13 | 9.7±1.9 |
| River bank | 28.3±2.7 | 8.75±0.23 | 12.2±3.6 | 24.6±2.3 | 8.68±0.17 | 9.3±2.6 | 18.3±1.8 | 8.63±0.15 | 6.4±1.8 |
Note: The data indicate mean value ± standard deviation, the sign “-” represents no values, and the sample time was June in 2018. |
Figure 5 Water sources and their proportions of the four desert vegetation species of the Ebinur Lake basin |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}