
Journal of Geographical Sciences >
Relationship of minimum winter temperature and temperature seasonality to the northern range limit and species richness of trees in North America
|
Hong Qian, E-mail: hong.qian@illinoisstatemuseum.org |
Received date: 2021-07-29
Accepted date: 2021-11-18
Online published: 2022-04-25
Supported by
National Key Research and Development Program(2019YFA0607302)
Biologists have considered both winter coldness and temperature seasonality as major determinants of the northern limits of plants and animals in the Northern Hemisphere, which in turn drive the well-known latitudinal diversity gradient. However, few studies have tested which of the two climate variables is the primary determinant. In this study, we assess whether winter coldness or temperature seasonality is more strongly associated with the northern latitudinal limits of tree species and with tree species richness in North America. Tree species were recorded in each of 1198 quadrats of 110 km × 110 km in North America. We used correlation and regression analyses to assess the relationship of the latitude of the northern boundary of each species, and of species richness per quadrat, with winter coldness and temperature seasonality. Species richness was analyzed within 38 longitudinal, i.e., north-south, bands (each being >1100 km long and 110 km wide). The latitudes of the northern range limits of tree species were three times better correlated with minimum temperatures at those latitudes than with temperature seasonality. On average, minimum temperature and temperature seasonality together explained 81.5% of the variation in the northern range limits of the tree species examined, and minimum temperature uniquely explained six-fold (33.7% versus 5.8%) more of this variation than did temperature seasonality. Correlations of tree species richness with minimum temperatures were stronger than correlations with temperature seasonality for most of the longitudinal bands analyzed. Compared to temperature seasonality, winter coldness is more strongly associated with species distributions at high latitudes, and is likely a more important driver of the latitudinal diversity gradient.
Hong QIAN , ZHANG Yangjian , Robert E. RICKLEFS , Xianli WANG . Relationship of minimum winter temperature and temperature seasonality to the northern range limit and species richness of trees in North America[J]. Journal of Geographical Sciences, 2022 , 32(2) : 280 -290 . DOI: 10.1007/s11442-022-1947-z
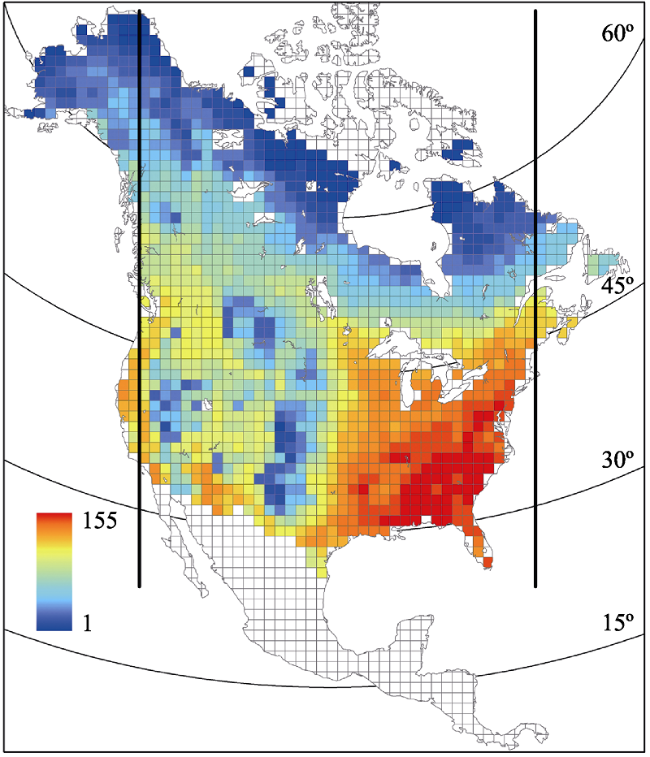
Figure 1 Geographic variation in tree species richness in North America north of Mexico. Each quadrat is 110 km by 110 km. Species richness in quadrats with land area less than 75% of a full quadrat is not shown. The 38 longitudinal bands between the two thick vertical lines were used in data analysis in this study. |
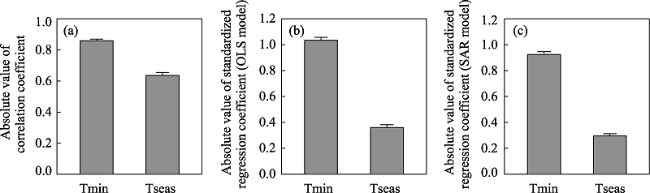
Figure 2 The means (± se) of (a) the absolute value of the correlation coefficient between northern latitude limit and either minimum temperature (Tmin) or temperature seasonality (Tseas), and absolute value of standardized partial regression coefficient of northern latitude limit against minimum temperature (Tmin) or temperature seasonality based on (b) ordinary least squares (OLS) and (c) simultaneous autoregression (SAR) models for 291 tree species in North America |
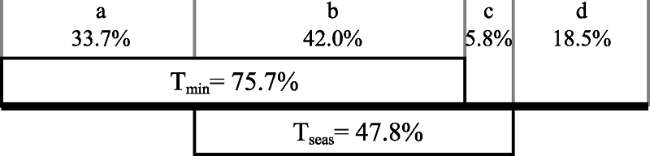
Figure 3 Results of a partial regression analysis (partitioning the variance). The variance in the response variable is partitioned into four portions: (a) uniquely accounted for by minimum temperature (Tmin), (b) accounted for jointly by minimum temperature and temperature seasonality (Tseas), (c) uniquely accounted for by temperature seasonality, and (d) unexplained variance. |
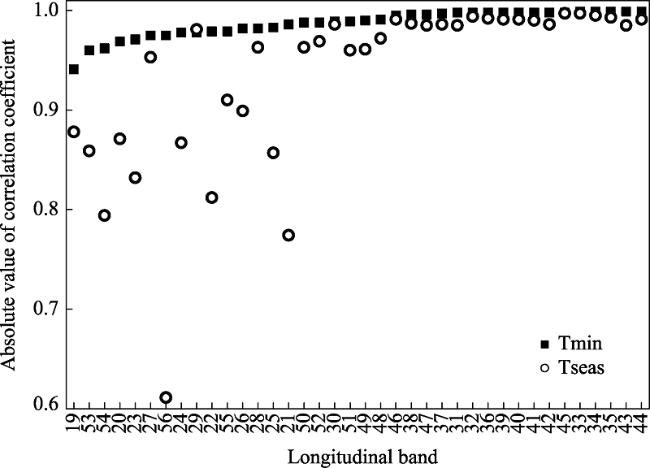
Figure 4 Comparison between the correlation between tree northern range limit and minimum temperature (Tmin) and between tree northern range limit and temperature seasonality (Tseas) for 38 longitudinal bands in North America (see Figure 1). Longitudinal bands were sorted according to absolute values of minimum temperature increasing from left to right. |
| [1] |
|
| [2] |
|
| [3] |
Behrensmeyer,
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}