
Journal of Geographical Sciences >
Dead shrub patches as ecosystem engineers in degraded drylands
|
Ilan STAVI, E-mail address: istavi@adssc.org |
Received date: 2020-12-31
Accepted date: 2021-03-16
Online published: 2021-10-25
A long-term drought has led to the mass mortality of shrubs in the semi-arid Israeli Negev. The most impacted shrub species is the Noaea mucronata (Forssk.) Asch. and Schweinf. In a four-year study, we found that herbaceous vegetation growth was greater in the dead shrub patches than in the surrounding inter-patch biocrusted spaces, suggesting that the dead shrub patches encompass improved micro-habitats. However, unexpectedly, the soil moisture in the dead shrub patches was consistently lower than that of the inter-patch biocrusted spaces. At the same time, soil quality in the dead shrub patches was higher than that in the inter-patch spaces. Therefore, it seems that the overall better soil conditions in the dead patches overcome the scarcity of soil-water, supporting increased herbaceous productivity. For explaining the discrepancy between herbaceous vegetation and soil-water, we formulated a conceptual framework, which highlights the key factors that regulate soil-water dynamics in this dryland ecosystem. We demonstrate that herbaceous vegetation is facilitated in the dead shrub patches by a legacy effect that takes place long after the shrubs have died. The dead shrub patches encompass a unique form of ecosystem engineering. The study high- lights the complex and unpredicted impacts of prolonged droughts on dryland ecosystems.
Ilan STAVI , Eli ZAADY , Alexander GUSAROV , Hezi YIZHAQ . Dead shrub patches as ecosystem engineers in degraded drylands[J]. Journal of Geographical Sciences, 2021 , 31(8) : 1187 -1204 . DOI: 10.1007/s11442-021-1892-2
Figure 1 Map of Israel, with an asterisk marking the study site |
Figure 2 A dead shrub patch, with a hammer at the patch's right edge.Note the considerably greater biomass of fresh herbaceous vegetation in the patch (the picture's center) than that in the inter-patch space (the picture's margins). Picture was taken in February 2018. |
Table 1 Microhabitat effect on soil properties |
| P value | Shrub patch | Inter-shrub space | |
|---|---|---|---|
| Clay (%) | 0.7412 | 16.0 a (0.4) | 16.2 a (0.6) |
| Silt (%) | 0.486 | 17.2 a (0.6) | 18.0 a (0.9) |
| Sand (%) | 0.3346 | 66.8 a (0.7) | 65.8 a (0.7) |
| Texture class | - | Sandy loam | Sandy loam |
| Bulk density (g/cm3) | < 0.0001 | 1.46 b (0.02) | 1.68 a (0.01) |
| Aggregate stability (%) | < 0.0001 | 69.9 a (2.4) | 39.9 b (1.6) |
| Clay dispersion index | < 0.0001 | 1.3 b (0.1) | 2.3 a (0.2) |
| Total organic carbon (g/kg) | < 0.0001 | 16.9 a (0.6) | 10.8 b (0.3) |
| Labile organic carbon (mg/kg) | < 0.0001 | 359.9 a (20.9) | 90.4 b (14.8) |
| Carbon lability (%/%) | < 0.0001 | 0.021301 a (0.000852) | 0.000773 b (0.001116) |
| N-NH4+ (mg kg-1) | < 0.0001 | 8.27 a (1.45) | 1.46 b (0.69) |
| N-NO2- (mg kg-1) | 0.5780 | 0.065 a (0.005) | 0.076 a (0.022) |
| Calcium carbonate (%) | 0.0196 | 12.2 b (0.2) | 13.1 a (0.3) |
| pH | < 0.0001 | 7.4 b (0.1) | 7.9 a (0.1) |
| Electrical conductivity (µS/cm) | < 0.0001 | 997.5 a (73.3) | 461.8 b (34.9) |
Notes: Bold P value indicates a significant effect. Means within the same row followed by a different letter differ at the 0.05 probability level according to Tukey's HSD. Numbers within parentheses are standard error of the means. |
Table 2 Depth effect on soil properties |
| P value | 0-5 cm | 5-10 cm | |
|---|---|---|---|
| Clay (%) | 0.4477 | 15.8 a (0.4) | 16.4 a (0.6) |
| Silt (%) | 0.3488 | 18.1 a (0.8) | 17.1 a (0.7) |
| Sand (%) | 0.6496 | 66.1 a (0.7) | 66.6 a (0.7) |
| Texture class | - | Sandy loam | Sandy loam |
| Bulk density (g/cm3) | 0.036 | 1.54 b (0.03) | 1.59 a (0.02) |
| Aggregate stability (%) | < 0.0001 | 62.4 a (3.4) | 47.5 b (2.6) |
| Clay dispersion index | 0.5072 | 1.7 a (0.2) | 1.9 a (0.2) |
| Total organic carbon (g/kg) | < 0.0001 | 15.4 a (0.8) | 12.3 b (0.5) |
| Labile organic carbon (mg/kg) | < 0.0001 | 284.0 a (31.7) | 166.4 b (25.7) |
| Carbon lability (%/%) | 0.0004 | 0.01679014 a (0.0014439) | 0.00773081 b (0.0016049) |
| N-NH4+ (mg kg-1) | < 0.0001 | 6.69 a (1.71) | 3.05 b (1.22) |
| N-NO2- (mg kg-1) | 0.0400 | 0.049 b (0.006) | 0.093 a (0.019) |
| Calcium carbonate (%) | 0.1709 | 12.4 a (0.2) | 12.9 a (0.3) |
| pH | < 0.0001 | 7.4 b (0.1) | 7.9 a (0.1) |
| Electrical conductivity (µS/cm) | < 0.0001 | 956.0 a (81.2) | 503.3 b (37.7) |
Notes: Bold P value indicates a significant effect. Means within the same row followed by a different letter differ at the 0.05 probability level according to Tukey's HSD. Numbers within parentheses are standard error of the means. |
Table 3 Effect of the interaction between microhabitat and depth on soil properties |
| Bulk density (g/cm3) | Aggregate stability (%) | Total organic carbon (g/kg) | Electrical conductivity (µS/cm) | |
|---|---|---|---|---|
| P value | 0.0154 | 0.0091 | 0.0033 | 0.001 |
| Shrub patch × 0-5 cm | 1.40 c (0.03) | 80.2 a (2.3) | 19.2 a (0.7) | 1310.5 a (86.2) |
| Shrub patch × 5-10 cm | 1.51 b (0.03) | 59.6 b (1.9) | 14.6 b (0.5) | 684.6 b (29.6) |
| Inter-shrub space x 0-5 cm | 1.68 a (0.01) | 44.6 c (2.5) | 11.6 c (0.4) | 601.6 b (44.0) |
| Inter-shrub space × 5-10 cm | 1.68 a (0.01) | 35.3 d (1.3) | 10.0 c (0.2) | 321.4 c (17.7) |
Notes: Bold P value indicates a significant effect. Means within the same column followed by a different letter differ at the 0.05 probability level according to Tukey's HSD. Numbers within parentheses are standard error of the means. |
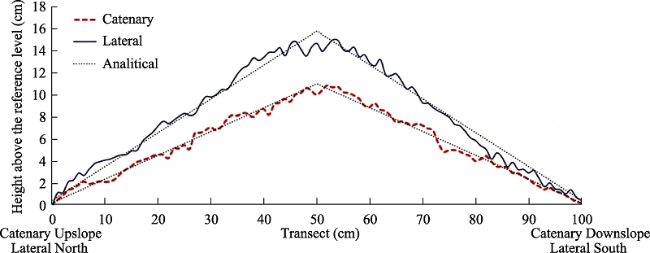
Figure 3 Microtopographic transects of mean catenary and lateral axes of dead shrub patches and their vicinities |
Table 4 Analytical model parameters for the catenary+lateral transects |
| Hmax (cm) | k | R2 | |
|---|---|---|---|
| Catenary transect | 11 | -0.22 | 0.888 |
| Lateral transect | 15.8 | -0.31 | 0.828 |
Notes: Hmax - the maximum mean value of transect height; k - transect slope coefficient; R2 - coefficient of determination. |
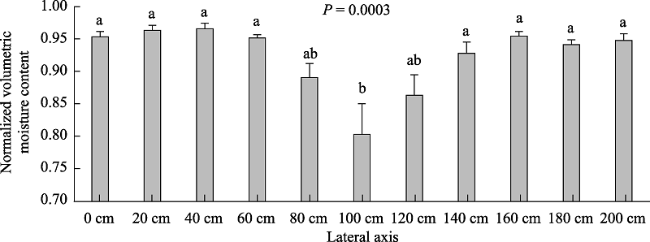
Figure 4 Mean normalized volumetric moisture content distribution along 2-m length lateral axis, during the rainy seasons of four consecutive years (2016/17 through 2019/20).Notes: the dead shrub's trunk is located in the axis' center (the 100 cm point). Error bars represent standard error of the means. Bars with different letters, differ at the 0.05 probability level according to Tukey's HSD. |
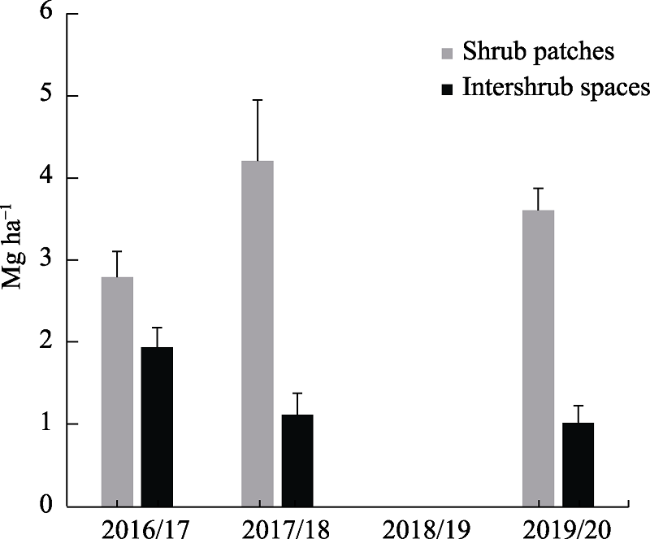
Figure 5 Mean herbaceous vegetation aboveground biomass in shrubby patches and adjacent intershrub spaces, according to growing season.Notes: error bars represent standard error of the means. |
Annex 1 Soil properties in the 0-10 cm depth of dead and live shrub patches, and the calculated ratio between them |
 |
Notes: Means of soil properties for dead shrub patches are combined values of the 0-5 and S-10 cm depths; data forsoil properties in the live shrub patches is obtained from Stavi et al. (2019). |
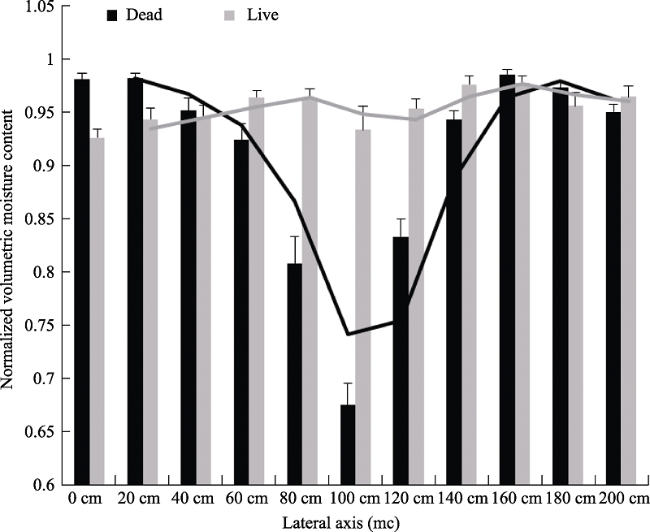
Annex 2 Mean normalized volumetric moisture content distribution and its moving average along 2-m length lateral axis of dead and live shrubs, during the rainy season.Note: Measurements were taken in nine dead and nine live shrub patches. Error bars represent standard error of the means. Lines represent moving averages. |
Figure 6 Schematic illustration of factors and processes that determine the soil-water dynamics in the shrubby patches and inter-patch spacesNotes: interception in the dead shrub patch is being conducted by the shrub canopy, the dense shoot of herbaceous plants, and the plant litter. In the inter-patch space, this process is conducted by the sparse herbaceous plants. Some interception may also be induced by the biological crusts. In the dead shrub patch, some of the raindrops may be accumulated in the upward-opening snail shells, where they remain until evaporated. Infiltration is facilitated in the dead shrub patch by the shrub root pathways, as well as by the dense root system of the herbaceous plants. In the inter-patch space, this process is facilitated by the sparse roots of herbaceous plants. Some infiltration may also be induced by the biological crusts. Transpiration is conducted in the dead shrub patch by the dense herbaceous plants. In the inter-patch space, this process is conducted by the sparse herbaceous plants. Some transpiration is also induced by the biological crusts. Evaporation is regulated in the dead shrub patch by the shrub canopy, plant litter, snail shells, and the dense herbaceous plants cover. In the inter-patch space, this process is regulated by the sparse herbaceous plants cover, as well as by the dense cover of biological crusts. Also, the biological crusts induce considerable dew condensation, of which some of it is uptaken by the crusts themselves. Despite the dense cover of plant litter and snail shells in the dead shrub patch, some runoff may be generated on its soil mound, down-mound flowing to the inter-patch space. In dead shrub patches, no hydraulic lift takes place. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
SAS Institute, 1990. SAS/STAT User's Guide, Version 6, 4th ed. SAS Inst., Cary, NC.
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
USDA-NRCS Dispersive clay soils. EFH NOTICE 210-WI-62. https://prod.nrcs.usda.gov/Internet/FSE_ DOCUMENTS/nrcs142p2_024606.pdf.
|
| [58] |
von Hardenberg J,
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}