
Journal of Geographical Sciences >
Biogeographical patterns of arbuscular mycorrhizal fungi diversity in China’s grasslands
|
Teng Jialing (1992-), PhD, specialized in microbial ecology. E-mail: tengjl92@gmail.com |
Received date: 2020-11-10
Accepted date: 2021-03-10
Online published: 2021-09-25
Supported by
National Key R&D Program of China(2017YFA0604803)
National Natural Science Foundation of China(31770560)
Copyright
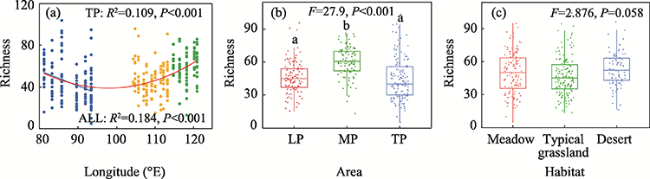
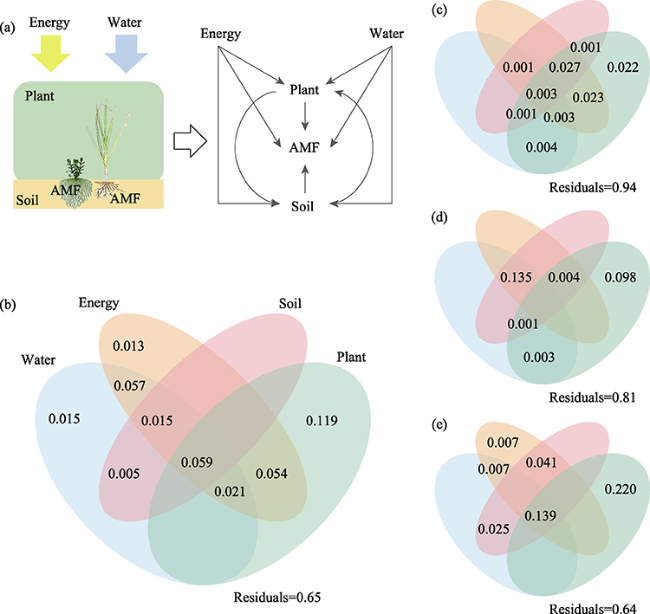
Arbuscular mycorrhizal fungi (AMF) are universally mutualistic symbionts that colonize the fine roots of most vascular plants. However, the biogeographical patterns and driving factors of AMF diversity of plant roots in grasslands are not well investigated. In this study, we used high-throughput sequencing techniques and bioinformatics to evaluate the AMF richness of 333 individual plant roots in 21 natural grassland ecosystems in northern China, including the Loess Plateau (LP), the Mongolian Plateau (MP), and the Tibetan Plateau (TP). The AMF richness showed a significant parabolic trend with increasing longitude. In regional situations, the AMF richness in the grasslands of the MP (60.4 ± 1.47) was significantly higher than those of the LP (46.4 ± 1.43) and TP (44.3 ± 1.64). Plant traits (including plant families, genera, and functional groups) explained the most variation in the AMF richness across China’s grasslands, followed by energy and water; soil properties had the least effects. The results showed the biogeographical patterns of the AMF richness and the underlying dominant factors, providing synthetic data compilation and analyses in the AMF diversity in China’s grasslands.
TENG Jialing , TIAN Jing , YU Guirui . Biogeographical patterns of arbuscular mycorrhizal fungi diversity in China’s grasslands[J]. Journal of Geographical Sciences, 2021 , 31(7) : 965 -976 . DOI: 10.1007/s11442-021-1880-6
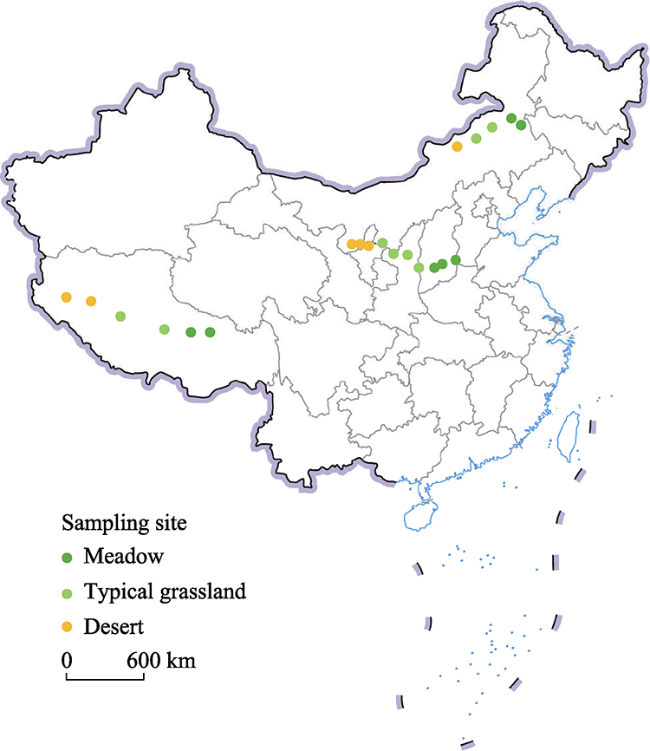
Figure 1 Geographic locations of sampling sites |
Table 1 List of environmental factors |
| Variables | Abbreviation | Unit | ||
|---|---|---|---|---|
| Climate | Energy | Mean annual temperature | MAT | ℃ |
| Mean temperature of coldest month | MTCM | ℃ | ||
| Mean temperature of warmest month | MTWM | ℃ | ||
| Annual total radiation | Rd | MJ m-2yr-1 | ||
| Maximum monthly radiation | Rdmax | MJ m-2day-1 | ||
| Minimum monthly radiation | Rdmin | MJ m-2day-1 | ||
| Minimum photosynthetically active radiation | PARmin | mol m-2day-1 | ||
| Maximum photosynthetically active radiation | PARmax | mol m-2day-1 | ||
| Potential evapotranspiration | ET0 | mm yr-1 | ||
| Water | Mean annual precipitation | MAP | mm | |
| Precipitation of the driest month | PDM | mm | ||
| Precipitation of the wettest month | PWM | mm | ||
| Other | Maximum monthly wind speed | Windmax | m s-1 | |
| Dryness index | ||||
| Soil | Organic carbon | SOC | % | |
| Carbon-Nitrogen ratio | CN | / | ||
| Total phosphorus | STP | mg kg-1 | ||
| Available phosphorus | SAP | mg kg-1 | ||
| NO3- | / | mg kg-1 | ||
| NH4+ | / | mg kg-1 | ||
| pH | / | / | ||
| Oxidation reduction potential | ORP | mv |
Figure 2 Longitudinal trends in arbuscular mycorrhizal fungi (AMF) richness (a) and AMF richness in different (b) areas and (c) habitats. Solid line represents model-predicted values of richness at a range of longitudes. Different letters indicate significant difference in richness between two groups. |
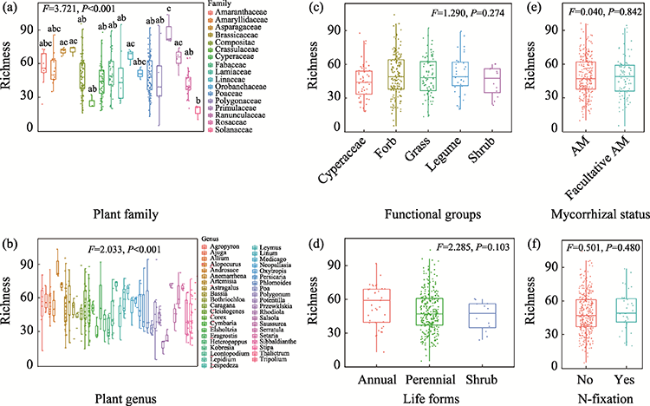
Figure 3 Arbuscular mycorrhizal fungi (AMF) richness of different plant traits (a. Families; b. Genera; c. Functional groups; d. Life forms; e. Mycorrhizal status; f. Nitrogen fixation. Different letters indicate significant difference in richness between two groups.) |
Table 2 Correlation coefficients and significance between arbuscular mycorrhizal fungi (AMF) richness and environmental variables |
| Variables | ALL | LP | MP | TP | |
|---|---|---|---|---|---|
| Energy | MAT | 0.052 | 0.074 | 0.245* | -0.118 |
| MTCM | -0.190** | 0.036 | 0.265* | -0.188* | |
| MTWM | 0.216** | 0.079 | 0.191 | 0.048 | |
| Rd | -0.155** | -0.063 | -0.181 | 0.299** | |
| Rdmax | 0.022 | -0.052 | -0.123 | 0.383** | |
| Rdmin | -0.298** | -0.093 | 0.13 | 0.01 | |
| PARmax | -0.069 | -0.069 | -0.207 | 0.341** | |
| PARmin | -0.297** | -0.099 | -0.015 | -0.052 | |
| ET0 | 0.196** | 0.241* | 0.245* | 0.189* | |
| Water | MAP | -0.213** | -0.094 | 0.16 | -0.352** |
| PDM | -0.203** | 0 | -0.201 | 0.003 | |
| PWM | -0.076 | -0.039 | 0.2 | -0.360** | |
| Other | Windmax | 0.255** | 0.200* | -0.011 | 0.244** |
| DI | -0.257** | -0.183 | -0.242* | -0.211* | |
| Soil | SOC | -0.160** | -0.027 | 0.128 | -0.225** |
| CN | -0.056 | -0.016 | 0.012 | 0.244** | |
| STP | -0.239** | -0.121 | -0.003 | -0.076 | |
| SAP | 0.052 | 0.101 | -0.344** | -0.157 | |
| NH4+ | -0.203** | -0.173 | -0.1 | -0.172* | |
| NO3- | -0.033 | -0.145 | -0.147 | -0.156 | |
| pH | 0.116* | -0.007 | 0.253* | 0.266** | |
| ORP | -0.211** | -0.099 | -0.015 | -0.052 |
Figure 4 Effects of water, energy, soil, and plant factors on the arbuscular mycorrhizal fungi (AMF) richness patterns (a. Diagrams showing biotic and abiotic factors affecting AMF richness; b. Entire grasslands; c. Loess Plateau (LP); d. Mongolian Plateau (MP); e. Tibetan Plateau (TP)) |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}