
Journal of Geographical Sciences >
North-south vegetation transition in the eastern Qinling-Daba Mountains
|
Zhang Xinghang (1992–), PhD Candidate, specialized in mountain geography and ecological environment. E-mail: jhj580724@l63.com |
Received date: 2020-10-15
Accepted date: 2020-12-21
Online published: 2021-05-25
Supported by
National Scientific and Technological Basic Resources Investigation Project(2017FY100900)
Copyright
The Qinling-Daba Mountains are the main body of China’s North-South Transitional Zone. Analysis of the north-south gradual variation of vegetation components is significant for understanding the structural diversity and complexity of this transitional zone. In this study, based on survey data of plant communities, the eastern Qinling-Daba Mountains is divided into four geographic units: the north flank of eastern Qinling Mts., south flank of eastern Qinling Mts., north flank of eastern Daba Mts. and south flank of eastern Daba Mts. We also explore division of regional climate according to areal differentiation of plant-species, community structure and species-richness, respectively. The results show that, (1) at plant-species level, there are mainly northern plants in north flank of eastern Qinling Mts. with evergreen species and fewer northern plants in south flank of eastern Qinling Mts.; there are mainly southern plants in eastern Daba Mts. (2) At community structure level, there are 4 formations (3 northern formations and 1 widespread formation) in north flank of eastern Qinling, 6 formations (3 northern formations, 1 southern formation, and 2 widespread formations) in south flank of eastern Qinling, 4 formations (2 southern formations and 2 widespread formations) in north flank of eastern Daba Mts., and 3 formations (3 southern formations) in south flank of eastern Daba Mts. In terms of the numbers and properties of formations, there is a mixture of northern and southern formations only in the south flank of eastern Qinling Mts. (3) At species-richness level, the diversity of families, genera and species decreased with increasing latitude, but the mixing of northern plants and the southern plants began to occur in south flank of eastern Qinling Mts. This means that the south flank of the eastern Qinling Mts. serves more suitably as the dividing line between China’s warm temperate and subtropical zones.
ZHANG Xinghang , ZHANG Baiping , WANG Jing , YU Fuqin , ZHAO Chao , YAO Yonghui . North-south vegetation transition in the eastern Qinling-Daba Mountains[J]. Journal of Geographical Sciences, 2021 , 31(3) : 350 -368 . DOI: 10.1007/s11442-021-1840-1
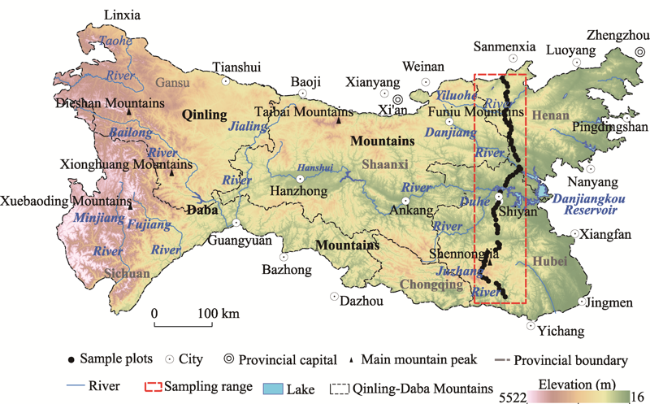
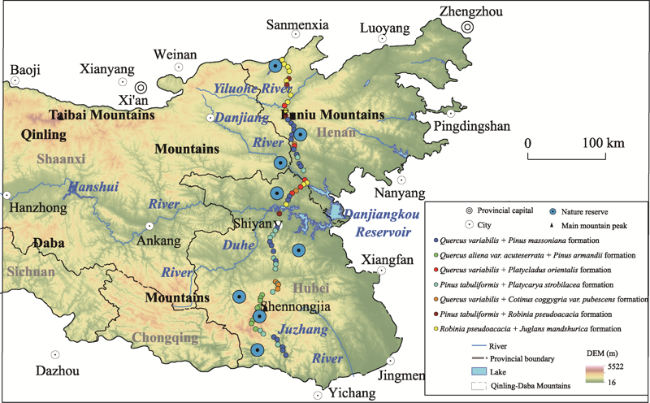
Figure 1 Sketch map of the Qinling-Daba Mountains and sampling sites |
Figure 2 Sequence of plant species along the north-south transect in the eastern Qinling-Daba Mountains |
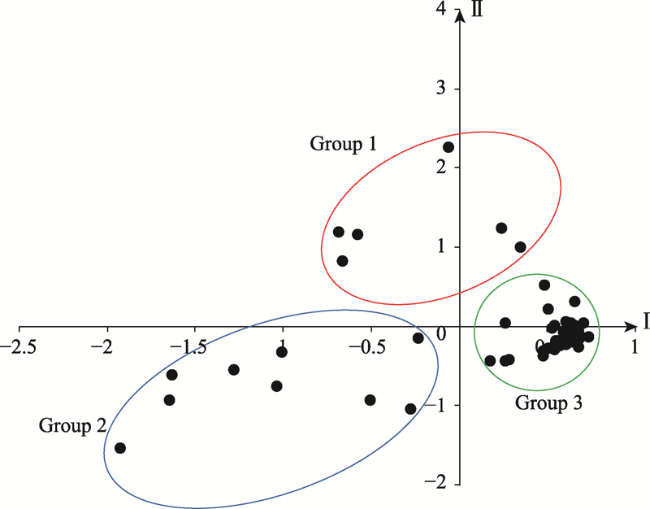
Table 1 Composition of plant properties based on the sorting results |
| Type | Feature | ||||
|---|---|---|---|---|---|
| Group | Transitional species | Widespread species | Concentrated distribution section | ||
| Northern plant type | Southern plant type | ||||
| Tree plants | 1 | 83% | 17% | 0 | Northern section |
| 2 | 0 | 78% | 22% | Southern section | |
| 3 | 13% | 0 | 87% | Northern and southern sections | |
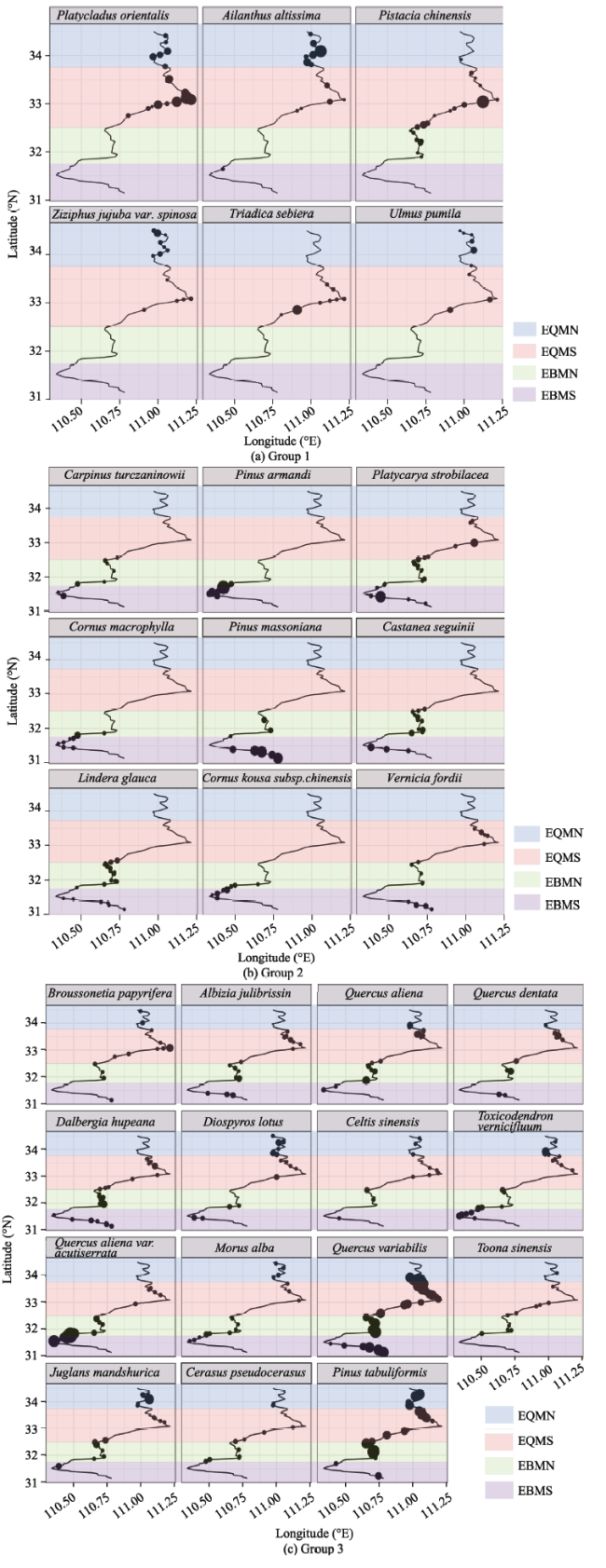
Figure 3 Bubble maps showing species abundance in the north-south direction. The bubble size is proportional to the current species abundance. |
Table 2 Distribution proportion of plant species in the north and south of the transect |
| Species in the north (25 species) | Species in the south (28 species) | ||
|---|---|---|---|
| Species found only in the north | Species found and distributed only in the south | Species found only in the south | Species found and distributed only in the north |
| 4 | 21 | 10 | 18 |
| 16% | 84% | 26% | 64% |
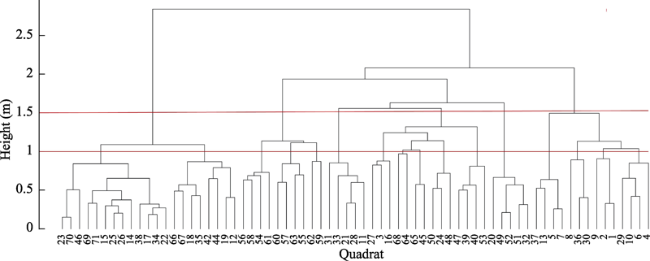
Figure 4 Ward cluster map of tree flora in the north-south transect in the eastern Qinling-Daba Mountains |
Figure 5 Spatial distribution of plant formations along the north-south transect of the eastern Qinling-Daba Mountains |
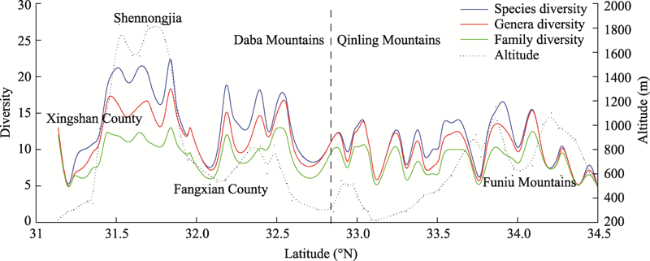
Figure 6 Latitudinal gradient changes in species richness along the north-south transect in the eastern Qinling-Daba Mountains |
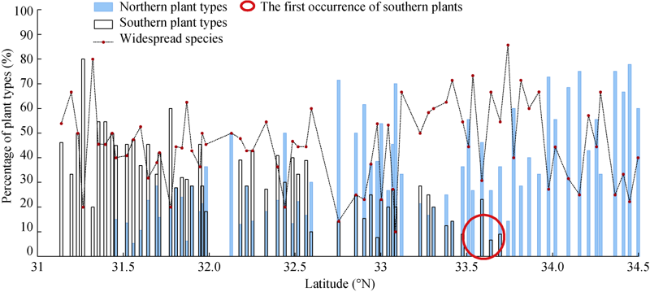
Figure 7 Latitudinal gradient changes in the species richness of different plant types along the north-south transect of the eastern Qinling-Daba Mountains |
Table 3 Basic data of some nature reserves along the north-south transect in the eastern Qinling-Daba Mountains |
| Natural Reserve | Position | Latitude (N) | Longitude (E) | Annual precipitation (mm) | Climate type | Belt types of mountain vegetation |
|---|---|---|---|---|---|---|
| Xiaoqinling | EQMN | 34°27′ | 110°33′ | 719.2 | Temperate and monsoonal climate | Low-mountain shrub meadow crop belt, deciduous oak forest belt |
| Baotianman | 33°30′ | 111°48′ | 885.6 | Temperate and monsoonal climate | Deciduous oak forest belt | |
| Xinkailing | EQMS | 33°16′ | 110°38′ | 803 | Subtropical and warm temperate monsoon climate | Evergreen and deciduous broad-leaved forest |
| Hualongshan | 33°07′ | 109°20′ | 1015 | Humid mountain climate in subtropical warm temperate zone | Broad-leaved evergreen forests | |
| Wudaoxia | EBMN | 32°05′ | 111°07′ | 1100 | North subtropical monsoon climate | Evergreen broad-leaved forest and deciduous broad-leaved mixed forest |
| Shibalichangxia | 31°31′ | 109°50′ | 1250 | North subtropical humid monsoon climate | Evergreen deciduous broad-leaved mixed forest | |
| Shennongjia | EBMS | 31°36′ | 110°27′ | 1200 | North subtropical monsoon climate | Broad-leaved evergreen forests |
| Wulipo | 31°22′ | 110°3′ | 1400 | Warm and moist monsoon climate in the middle subtropics | Evergreen broad-leaved forest and evergreen coniferous forest belt |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}