
Journal of Geographical Sciences >
Spatial patch structure and adaptive strategy for desert shrub of Reaumuria soongorica in arid ecosystem of the Heihe River Basin
|
Li Wei (1987–), PhD, specialized in ecohydrological processes and water resource management. E-mail: liwei19870316@hotmail.com |
Received date: 2019-01-30
Accepted date: 2019-03-30
Online published: 2019-12-11
Supported by
National Natural Science Foundation of China(No.91425301)
National Natural Science Foundation of China, No.91425301(No.41730854)
The PCSIRT(No.IRT-15R06)
Scientific Research Starting Foundation for Doctors in Hebei GEO University(No.BQ2017003)
Copyright
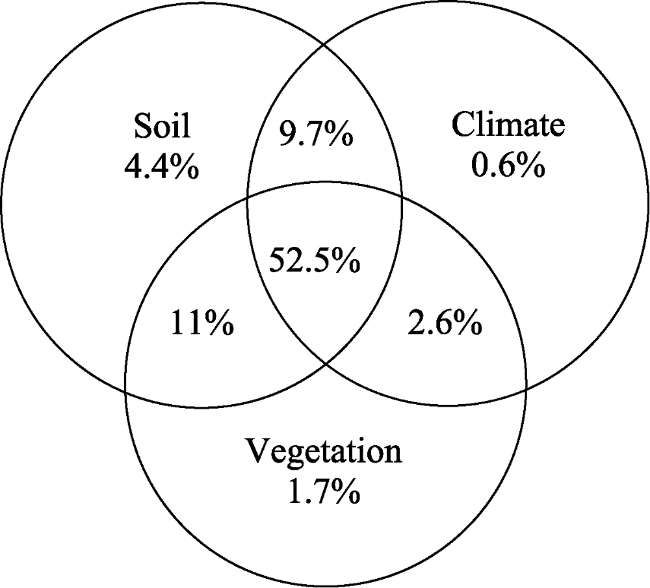
In many arid ecosystems, vegetation frequently occurs in high-cover patches interspersed in a matrix of low plant cover. However, theoretical explanations for shrub patch pattern dynamics along climate gradients remain unclear on a large scale. This context aimed to assess the variance of the Reaumuria soongorica patch structure along the precipitation gradient and the factors that affect patch structure formation in the middle and lower Heihe River Basin (HRB). Field investigations on vegetation patterns and heterogeneity in soil properties were conducted during 2014 and 2015. The results showed that patch height, size and plant-to-patch distance were smaller in high precipitation habitats than in low precipitation sites. Climate, soil and vegetation explained 82.5% of the variance in patch structure. Spatially, R. soongorica shifted from a clumped to a random pattern on the landscape towards the MAP gradient, and heterogeneity in the surface soil properties (the ratio of biological soil crust (BSC) to bare gravels (BG)) determined the R. soongorica population distribution pattern in the middle and lower HRB. A conceptual model, which integrated water availability and plant facilitation and competition effects, was revealed that R. soongorica changed from a flexible water use strategy in high precipitation regions to a consistent water use strategy in low precipitation areas. Our study provides a comprehensive quantification of the variance in shrub patch structure along a precipitation gradient and may improve our understanding of vegetation pattern dynamics in the Gobi Desert under future climate change.
LI Wei , LI Xiaoyan , HUANG Yongmei , WANG Pei , ZHANG Cicheng . Spatial patch structure and adaptive strategy for desert shrub of Reaumuria soongorica in arid ecosystem of the Heihe River Basin[J]. Journal of Geographical Sciences, 2019 , 29(9) : 1507 -1526 . DOI: 10.1007/s11442-019-1674-2
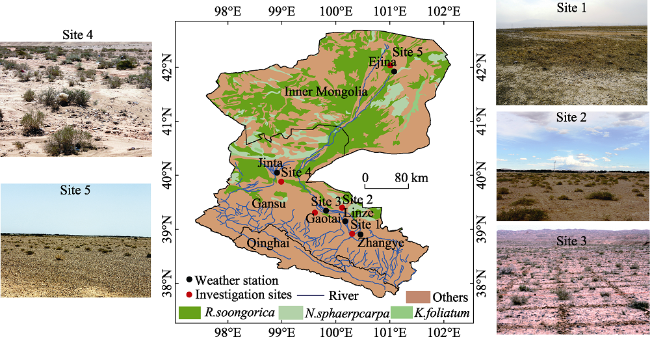
Figure 1 Location of the Heihe River Basin: field investigation sites (red dots) and corresponding weather stations (black dots); others are images of the landscape and typical shrubs at the study sites |
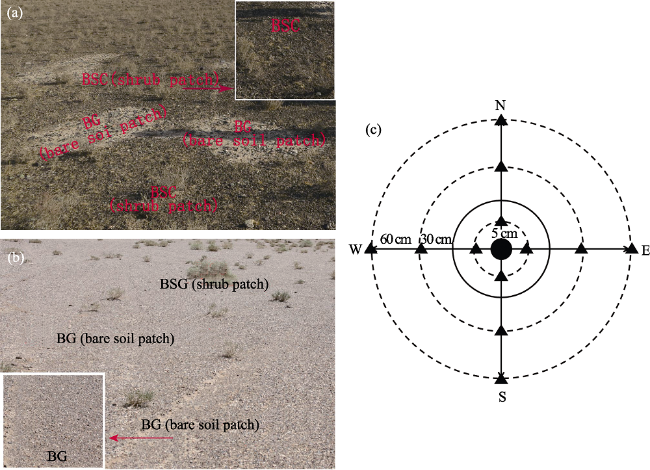
Figure 2 Spatial heterogeneity in surface soil properties, consisting of biological soil crust (BSC) developed with shrub patch and bare soil patch covered with fine gravels (BG) at (a) Site 1 and (b) Site 5; (c) surface soil sampling design for individual shrubs, the bold rings represent the average extent of shrub size |
Table 1 Summary of climate and soil properties in the middle and lower Heihe River Basin |
| Site 1 Piedmont Gobi desert | Site 2 Piedmont Gobi desert | Site 3 Piedmont Gobi desert | Site 4 Gobi desert | Site 5 Gobi desert | |
|---|---|---|---|---|---|
| Longitude | 100.30°E | 100.12°E | 99.61°E | 98.99°E | 101.01°E |
| Latitude | 38.91°N | 39.40°N | 39.31°N | 39.88°N | 42.04°N |
| Elevation (m) | 1473 | 1442 | 1355 | 1413 | 940 |
| Main shrub | R.soongorica; S.passerine | R.soongorica; N.sphaerpcarpa | R.soongorica; K.foliatum | R.soongorica; N.sphaerpcarpa | R.soongorica; H.ammodendron |
| MAP (mm) | 122.93a | 112.29b | 100.33b | 65.70c | 35.02d |
| MAT (℃) | 7.32a | 7.99ab | 7.87bc | 9.02d | 9.11e |
| Sand content (%) | 57.65 | 89.85 | 86.82 | 89.44 | 97.51 |
| Silt content (%) | 39.46 | 8.13 | 11.50 | 8.75 | 1.68 |
| Clay content (%) | 2.89 | 2.02 | 1.68 | 1.81 | 0.81 |
| SOM content (%) | 1.97 | 0.97 | 0.73 | 1.84 | 0.62 |
| TN content (%) | 0.09 | 0.02 | 0.05 | 0.02 | 0.01 |
| CaCO3 content (%) | 44.12 | 30.71 | 14.64 | 42.57 | 35.61 |
| BSC:BG | 0.13:1 | 0.09:1 | 0.06:1 | 0.06:1 | 0.02:1 |
Note: Different letters donate different means at 95% confidence level. MAP is mean annual precipitation (mm); MAT is mean annual temperature (℃); soil particles including sand, silt and clay content is defined in 0-10 cm depth; SOM is soil organic matter in 0-10 cm depth; TN is total N in 0-10 cm depth and CaCO3 is defined in 0-10 cm depth; BSC:BG is the ratio of biological soil crust and gravel content at soil surface. |
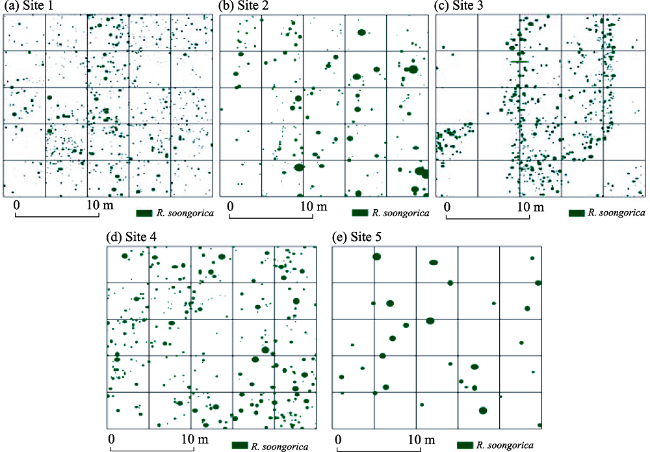
Figure 3 Shrub patch distributions (25 m × 25 m) of R. soongorica from Site 1 to Site 5 in the middle and lower Heihe River Basin |
Table 2 Summary of R. soongorica patch structure across the mean annual precipitation gradient |
| Site | Percentage (%) | N | C | KA (m2) | H (cm) | $\bar{A}$(m2) |
|---|---|---|---|---|---|---|
| Site 1 | 63.4a | 1049±128a | 1.68±0.19a | 4.1±0.7a | 10.15±0.32a | 0.02±0.01a |
| Site 2 | 58.5a | 174±80ab | 0.33±0.14b | 2.6±1.0a | 18.08±0.31b | 0.10±0.02ab |
| Site 3 | 89.4b | 821±261a | 1.08±0.40a | 4.6±0.4b | 17.53±2.36bc | 0.04±0.01a |
| Site 4 | 92.6b | 356b | 0.57±0.00b | 4.7±0.2b | 17.57±0.34bc | 0.08±0.00ab |
| Site 5 | 95.1b | 48±11c | 0.08±0.01c | 1.6±1.0c | 23.10±5.60d | 0.21±0.11b |
Note: Different letters donate different means at 95% confidence level. Percent is ratio of number of R.soongorica to total plants number (%); N: total number of shrub patches; C: density of shrub patches per m-2; KA: total area of shrub patches (%); H is the maximal height of shrub (cm); $\bar{A}$: size of each shrub patch (m2) |
Table 3 Summary statistics and statistical differences of vegetation community characteristics |
| Site | Cover (%) | AGB (g/m2) | SR | Simpson index | Shannon index | Important value | V/m | PD (m) |
|---|---|---|---|---|---|---|---|---|
| Site 1 | 11.16±0.03a | 92.36±15.21a | 4 | 0.69 | 1.28 | 0.42 | 13.4 | 0.56±0.04a |
| Site 2 | 8.03±0.04b | 38.64±10.29b | 3 | 0.64 | 1.05 | 0.42 | 2.9 | 1.40±0.33c |
| Site 3 | 5.33±0.01c | 25.92±7.35c | 3 | 0.61 | 1.06 | 0.58 | 5.3 | 0.60±0.03a |
| Site 4 | 5.61±0.00c | 27.01±0.17c | 2.5 | 0.50 | 0.74 | 0.66 | 3.4 | 0.95±0.00b |
| Site 5 | 1.85±0.01d | 8.14±1.20d | 2 | 0.51 | 0.63 | 0.68 | 0.9 | 2.80±0.54d |
Note: Different letters donate different means at 95% confidence level. AGB is above-ground biomass; SR is species richness; important value is related to R.soongorica shrub; PD is patch-to-patch distance (m). |
Table 1S Pearson’s correlation analysis between shrub patch structure index and environmental factors (climate, soil and vegetation) |
| Factors structure | Climate | Soil | Vegetation | |||
|---|---|---|---|---|---|---|
| MAP | Sand | TN | BSC:BG | AGB | RS | |
| Height | -0.71** | 0.80** | -0.74** | -0.67** | -0.78** | -0.76** |
| Size | -0.69** | 0.62** | -0.54** | -0.51* | -0.53* | -0.58* |
| Cover | 0.85** | -0.85** | 0.72** | 0.99** | 0.91** | 0.80** |
| Density | 0.73** | -0.87** | 0.95** | 0.77** | 0.87** | 0.83** |
| PD | -0.79** | 0.69** | -0.61** | -0.68** | -0.61** | -0.61** |
| V/m | 0.76** | -0.93** | 0.93** | 0.77** | 0.90** | 083** |
*Correlations are significant at P < 0.05; **Correlations are significant at P < 0.01 |
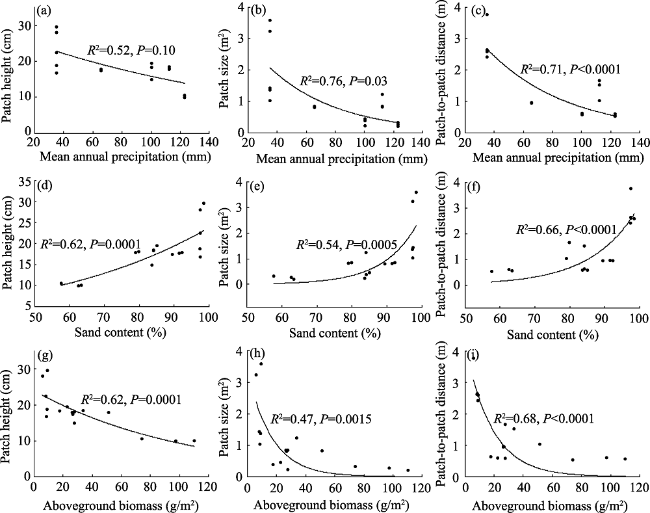
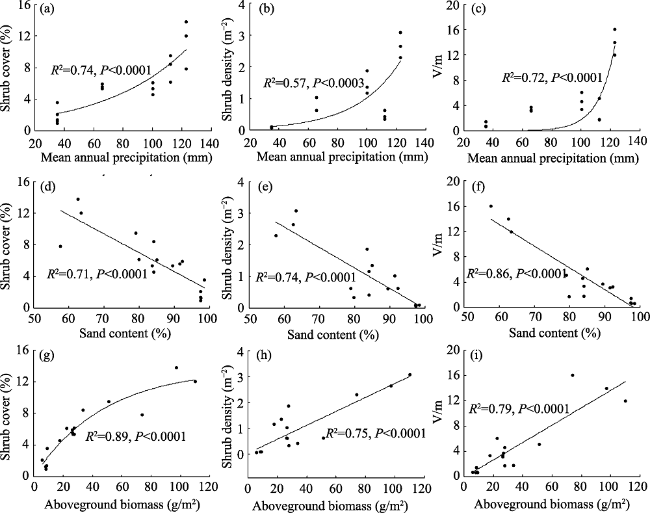
Figure 4 Changes in shrub patch attributes with (a-c) mean annual precipitation (MAP), (d-f) clay content of 0-10 cm, and (g-i) community aboveground biomass (ABG). The shrub patch structure includes mean patch height, size and plant distance. |
Changes in community characteristics with (a-c) mean annual precipitation, (d-f) clay content of 0-10 cm, (g-i) community aboveground biomass. The community characteristics included vegetation cover, density and variance to mean ratio (V/m). |
Table 4 Regression analysis between R. soongorica patch structure patch and climate, soil and vegetation factors |
| Dependent factor | Independent factor | Regression function |
|---|---|---|
| Height | Sand content | y=0.33x-0.97 R2=0.64 P<0.001 |
| Size | MAP | y=-0.002x-0.26 R2=0.44 P<0.01 |
| Distance | MAP | y=-0.02x+3.23 R2=0.61 P<0.001 |
| Shrub cover | MAP, species richness, BSC:BG | y=0.007x1-0.19x2+84.70x3+0.46 R2=0.99 P<0.001 |
| Shrub density | TN | y=34.48x-0.03 R2=0.89 P<0.001 |
| V/m | MAP, sand content | y=-0.07x1-0.52x2+54.60 R2=0.93 P<0.001 |
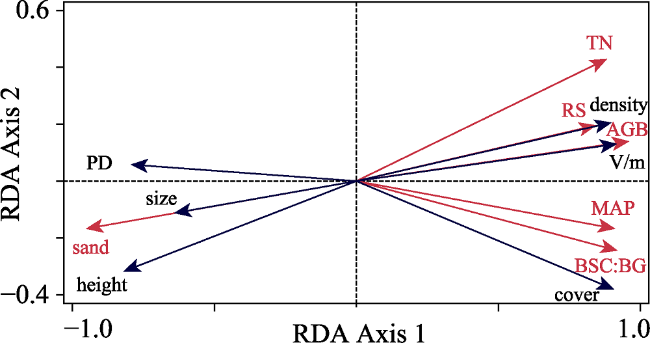
Figure 6 Redundancy analysis (RDA) displays the effects of climate, soil, and vegetation index indicators (red vectors) on shrub patch structure variables (blue vectors) |
Figure 7 Combined effects of MAP, soil properties and vegetation on shrub patch structure |
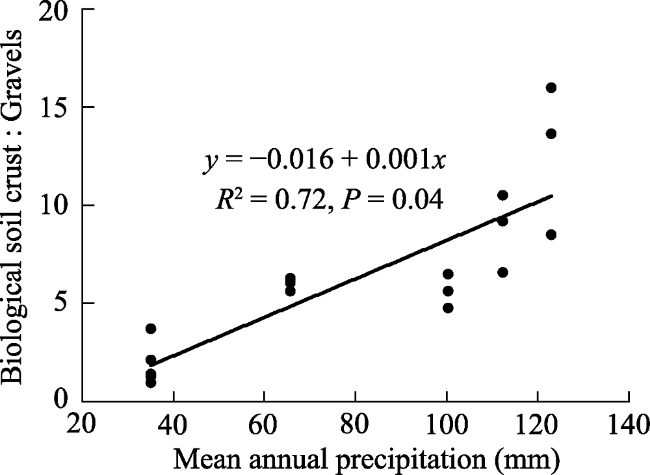
Figure 8 Relationship between MAP and surface soil heterogeneity (the ratio of biological soil crust and bare gravel) |
Table 5 Pearson correlation between the ratio of biological soil crust (BSC) and bare gravel (BG) and shrub patch structure and spatial pattern |
| H | $\bar{P}$ | $\bar{A}$ | PD | SR | AGB | IV | |
|---|---|---|---|---|---|---|---|
| BSC:BG | -0.97** | -0.78 | -0.75 | -0.70 | 0.95* | 0.95* | -0.88* |
| BSC:BG (MAP) | -0.16 | 0.26 | 0.24 | -0.01 | 0.66** | 0.78** | -0.34 |
Note: *Correlations are significant at P < 0.05; **Correlations are significant at P < 0.01 |
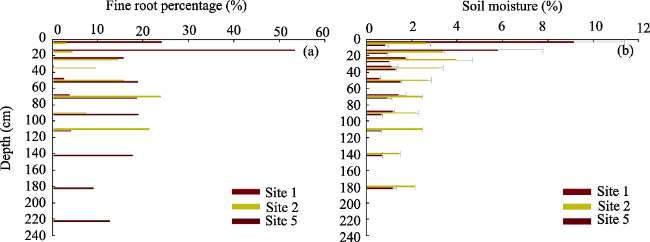
Figure 1S A comparison of vertical pattern of surface area of fine root (< 2 mm) and soil moisture in growing season of R. soongorica |
Table 2S Comparison of soil moisture, soil organic matter, total nitrogen and particle size from biological soil crust (BSC) and bare gravels (BG) |
| Sites | Type | SM | SOM | TN | Soil particles | ||
|---|---|---|---|---|---|---|---|
| Sand | Loam | Clay | |||||
| Site 1 | BSC | 7.88±1.14% | 4.28±.97% | 0.14±0.11% | 64.95% | 31.30% | 3.75% |
| BG | 5.44±1.09% | 3.83±0.58% | 0.07±0.03% | 83.52% | 10.43% | 6.05% | |
| FIE | 1.44 | 1.12 | 1.85 | 0.78 | 3.00 | 0.62 | |
| Site 2 | BSC | 3.24±1.27% | 2.13±0.30% | 0.02±0.00% | 79.43% | 18.15% | 2.42% |
| BG | 2.53±1.00% | 2.13±0.46% | 0.02±0.01% | 88.48% | 9.42% | 2.10% | |
| FIE | 1.27 | 1 | 1 | 0.90 | 1.93 | 1.15 | |
| Site 5 | BSC | 0.88±0.35% | 1.27±0.30% | 0.01±0.00% | 97.00% | 1.75% | 1.25% |
| BG | 0.95±0.54% | 1.33±0.41% | 0.01±0.00% | 97.90% | 1.28% | 0.82% | |
| FIE | 0.93 | 0.96 | 1 | 0.99 | 1.36 | 1.52 | |
Note: SM: soil moisture; FIE: “fertile islands” effects |
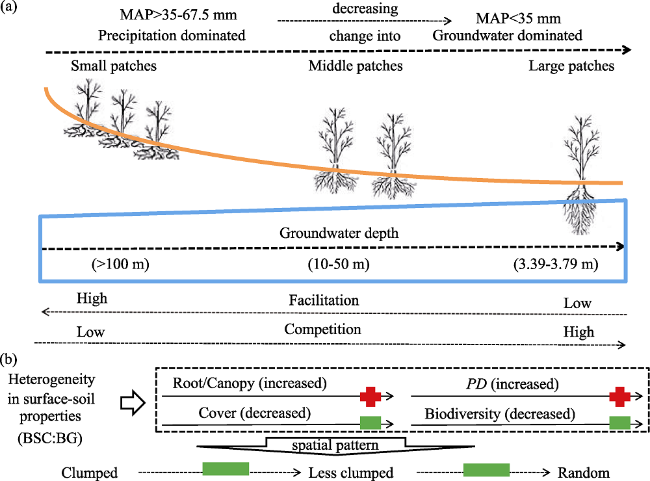
Figure 9 A conceptual model of patchy vegetation dynamics and ecological adaptive strategies of R. soongorica in the middle and lower Heihe River Basin |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}