
Journal of Geographical Sciences >
Moisture content variations in soil and plant of post-fire regenerating forests in central Yunnan Plateau, Southwest China
*Corresponding author: Shen Zehao, Professor, specialized in landscape ecology. E-mail: shzh@pku.edu.cn
Author: Chen Lifang, graduate student of landscape ecology. E-mail: chenlf0915@pku.edu.cn
Received date: 2018-05-10
Accepted date: 2019-01-22
Online published: 2019-07-25
Supported by
National Natural Science Foundation of China, No.41371190
Copyright
Plant moisture content (PMC) is used as an indicator of forest flammability, which is assumed to be affected by climate drought. However, the fire-induced drought stress on PMC and its spatial and temporal variations are unclear. Based on a parallel monitoring experiment from 2014 to 2015, this study compared the PMCs and soil moisture contents (SMC) at five post-fire sites in central Yunnan Plateau, Southwest China. The number of years since last fire (YSF), season, topographic position, plant species and tissue type (leaf and branch) were selected as causal factors of the variations in PMC and SMC. A whole year parallel monitoring and sampling in the post-fire communities of 1, 2, 5, 11 and 30 YSF indicated that drought stress in surface soils was the strongest in spring within the first 5 years after burning, and the SMC was regulated by topography, with 64.6% variation in soil moisture accounted for by YSF (25.7%), slope position (22.1%) and season (10.8%). The temporal variations of PMC and SMC differed at both interannual and seasonal scales, but the patterns were consistent across topographic positions. PMC differed significantly between leaves and branches, and among three growth-forms. The mean PMC was lower in broad-leaved evergreen species and higher in conifer species. Season and soil temperature were the primary determinants of PMC, accounting for 19.1% and 8.3% of variation in PMC, respectively. This indicated phenology-related growth rather than drought stress in soil as the primary driver of seasonal changes in PMC. The significant variations of PMC among growth forms and species revealed that seasonal soil temperature change and dominant species in forest communities are useful indicators of fire risk assessment in this region.
CHEN Lifang , DOU Qun , ZHANG Zhiming , SHEN Zehao . Moisture content variations in soil and plant of post-fire regenerating forests in central Yunnan Plateau, Southwest China[J]. Journal of Geographical Sciences, 2019 , 29(7) : 1179 -1192 . DOI: 10.1007/s11442-019-1652-8
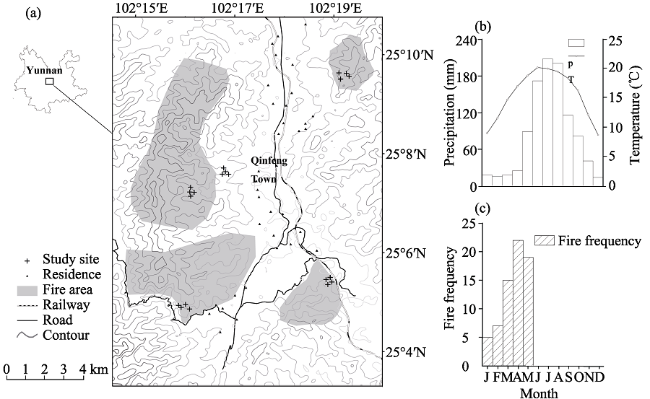
Figure 1 Geographical location and climatic features of the burned sites in central Yunnan Plateau: a) Location of the study area and the burned sites of 1, 2, 5, 11 and 30 years-since-last-fire (YSF); b) monthly mean precipitation and temperature in Qinfeng County; and c) monthly frequency distribution of fire records from 2003-2012 |
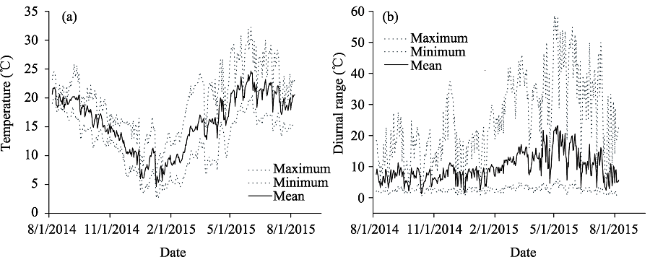
Figure 2 Temporal (a) and diurnal (b) dynamics of soil surface temperature, pooled for records collected on different slope positions of the five study sites in central Yunnan Plateau |
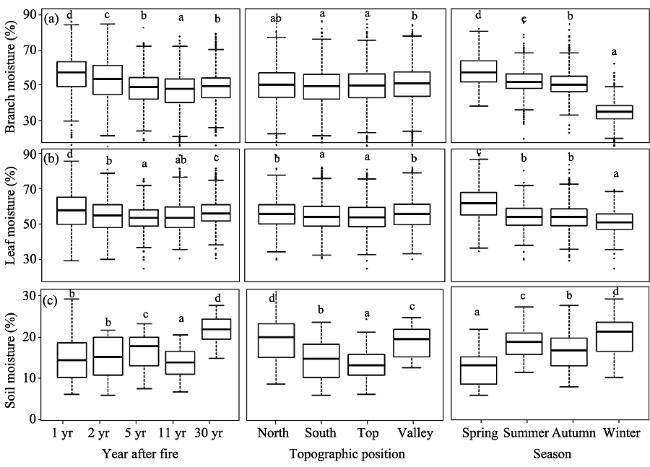
Figure 3 Plant branches/leaves moisture and soil moisture variation tendency at spatial and temporal scales in central Yunnan Plateau |
Table 1 The variations of moisture contents in the branches and leaves of trees in three growth forms with regard to YSF, season, and topographic position, in central Yunnan Plateau. Superscript character indicates the difference among values for different levels of each predictive factor. DB, deciduous broad-leaved species; EB, evergreen broad-leaved species; EC, evergreen coniferous species; South, south slope; North, north slope. |
| Plant tissue | Branch (%) | Leaf (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| Growth form | DB | EB | EC | DB | EB | EC | ||
| YSF | 1st | 60.8b±8.5 | 57.7d±8.8 | 61.3cd±7.9 | 60.0c±8.2 | 59.0b±8.5 | 66.3d±8.1 | |
| 2nd | 57.7b±8.3 | 53.3c±8.5 | 61.7d±8.9 | 59.1b±8.2 | 54.7a±8.6 | 63.6c±8.5 | ||
| 5th | 50.5a±7.7 | 49.5b±7.3 | 57.9bc±7.8 | 54.6a±7.6 | 55.4a±7.5 | 57.2b±7.4 | ||
| 11th | 50.0a±8.1 | 47.5a±6.9 | 56.7ab±8.5 | 57.9b±8.9 | 55.2a±7.1 | 59.8a±7.9 | ||
| 30th | 51.6a±7.8 | 50.6b±7.7 | 54.3a±7.9 | 57.0b±7.3 | 58.9b±8.1 | 60.8a±6.9 | ||
| Season | Spring | 62.1d±8.5 | 59.7c±8.8 | 67.7c±7.9 | 67.3c±8.2 | 62.9c±8.5 | 62.7c±8.1 | |
| Summer | 56.2c±8.2 | 54.0b±8.1 | 60.1b±7.8 | 55.3b±7.4 | 55.1b±7.7 | 61.1b±76 | ||
| Autumn | 53.1b±8.2 | 53.4b±7.8 | 59.6b±8.3 | 54.0b±8.8 | 55.8b±7.2 | 61.3b±8.3 | ||
| Winter | 38.3a±7.4 | 36.6a±7.0 | 40.1a±7.0 | 51.3a±7.2 | 52.4a±7.0 | 58.1a±7.5 | ||
| Topographic feature | North | 52.9a±8.5 | 51.8bc±8.8 | 58.3a±7.9 | 57.2a±8.2 | 57.3b±8.5 | 61.8b±8.1 | |
| South | 53.5a±8.9 | 49.6a±8.4 | 57.9a±8.3 | 58.1a±9.1 | 55.2a±8.7 | 60.8ab±8.3 | ||
| Top | 53.3a±8.4 | 50.6ab±8.6 | 57.0a±8.1 | 57.5a±8.2 | 55.0a±8.4 | 60.2a±8.6 | ||
| Valley | 53.7a±8.6 | 52.7c±8.7 | 59.4a±8.4 | 56.8a±8.3 | 58.2b±8.8 | 61.3ab±8.7 | ||
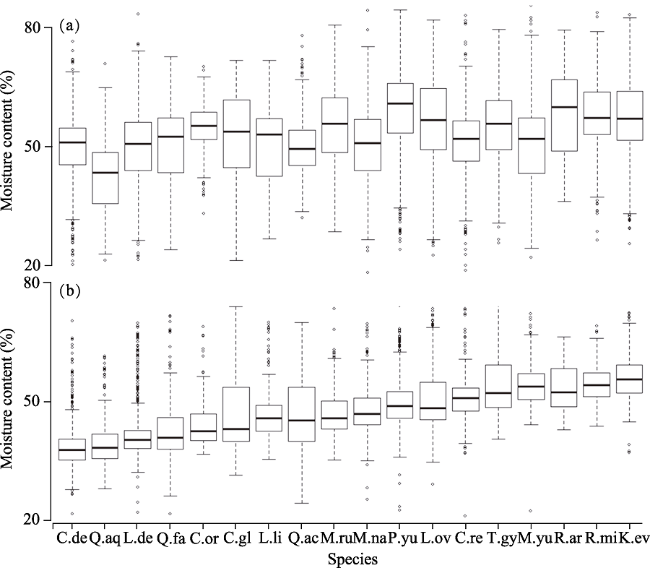
Figure 4 Branch (a) and leaf (b) moisture content of 18 plant species in central Yunnan Plateau. C.re, Camellia reticulata; C.or, Castanopsis orthacantha; C.de, Cyclobalanopsis delavayi; C.gl, C. glaucoides; K.ev, K. evelyniana; L.de, L. dealbatus; L.li, L. litseifolius; L.ov, Lyonia ovalifolia; M.yu, Michelia yunnanensis; M.na, M. nana; M.ru, M. rubra; P.yu, P. yunnanensis; Q.fa, Q. fabrei; Q.aq, Q. aquifolioides; Q.ac, Q. acutissima; R.mi, Rhododendron microphyton; R.ar, R. araiophyllum; T.gy, Ternstroemia gymnanthera |
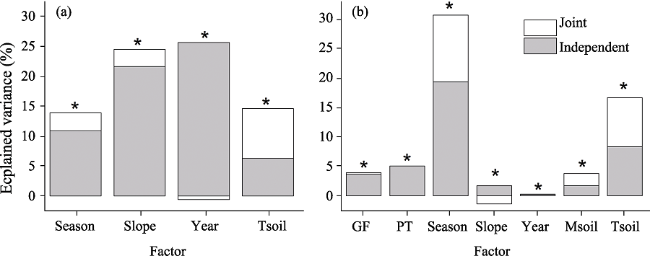
Figure 5 Hierarchical partition of moisture variation in (a) soil and (b) post-fire regenerated plant tissues in central Yunnan Plateau, based on GLM analysis (type includes three growth-forms covering DB, EB and EC, as indicated in the text, * means difference is statistically significant). |
Table 2 Hierarchical variation partitioning for PMC of three growth-forms and two types of plant tissue in central Yunnan Plateau. DB, deciduous broad-leaved; EB, evergreen broad-leaved; EC, evergreen coniferous. Indep., independent effect (%); Joint, joint effect (%). SMC, soil moisture content; Tsoil, temperature of surface soil. |
| Exp. variable | DB | EB | EC | Branch | Leaf | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Indep. | Joint | Indep. | Joint | Indep. | Joint | Indep. | Joint | Indep. | Joint | |
| Plant tissue | 3.74 | 0.00 | 6.85 | 0.00 | 2.29 | 0.00 | — | — | — | — |
| Growth-form | — | — | — | — | — | — | 3.55 | 0.06 | 4.35 | 0.40 |
| Season | 25.91 | 15.89 | 19.57 | 9.99 | 15.51 | 11.92 | 13.30 | 3.86 | 27.93 | 20.19 |
| Slope position | 1.17 | -1.09 | 2.60 | -1.15 | 1.82 | -1.37 | 1.24 | -0.60 | 2.57 | -2.33 |
| YSF | 0.69 | 0.49 | 0.17 | -0.16 | 1.17 | 1.18 | 0.52 | -0.26 | 1.12 | 0.73 |
| SMC | 3.88 | 5.58 | 1.23 | 0.63 | 2.43 | 3.29 | 0.81 | 0.18 | 3.10 | 4.62 |
| Tsoil | 11.35 | 12.78 | 7.48 | 6.46 | 9.82 | 9.15 | 2.76 | 1.85 | 17.49 | 15.95 |
The authors have declared that no competing interests exist.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|
| 54 |
|
| 55 |
|
| 56 |
|
| 57 |
|
| 58 |
|
| 59 |
|
| 60 |
|
| 61 |
|
| 62 |
|
| 63 |
|
| 64 |
|
| 65 |
|
| 66 |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}