
Journal of Geographical Sciences >
Plant functional traits and microbes vary with position on striped periglacial patterned ground at Glacier National Park, Montana
Author: APPLE Martha Elizabeth, E-mail: MApple@mtech.edu
Received date: 2018-05-10
Accepted date: 2019-01-22
Online published: 2019-07-25
Copyright
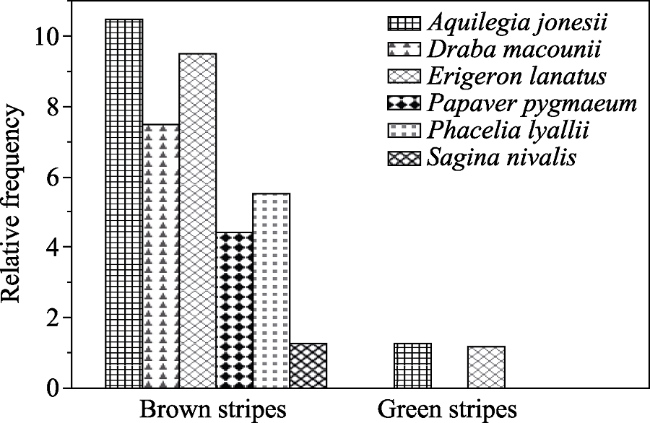
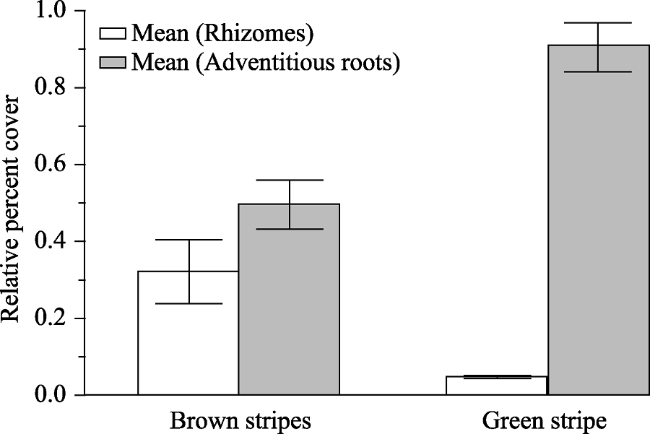
The retreating snowfields and glaciers of Glacier National Park, Montana, USA, present alpine plants with changes in habitat and hydrology. The adjacent and relic periglacial patterned ground consists of solifluction terraces of green, vegetation-rich stripes alternating with sparsely vegetated brown stripes. We established georeferenced transects on striped periglacial patterned ground for long-term monitoring and data collection on species distribution and plant functional traits at Siyeh Pass and at Piegan Pass at Glacier National Park. We documented species distribution and calculated the relative percent cover (RPC) of qualitative functional traits and used 16S rRNA from soil samples to characterize microbial distribution on green and brown stripes. Plant species distribution varied significantly and there were key differences in microbial distribution between the green and brown stripes. The rare arctic-alpine plants Draba macounii, Papaver pygmaeum, and Sagina nivalis were restricted to brown stripes, where the RPC of xeromorphic taprooted species was significantly higher at the leading edge of the Siyeh Pass snowfield. Brown stripes had a higher percentage of the thermophilic bacteria Thermacetogenium and Thermoflavimicrobium. Green stripes were co-dominated by the adventitiously-rooted dwarf shrubs Salix arctica and the possibly N-fixing Dryas octopetala. Green stripes were inhabited by Krummholz and seedlings of Abies lasiocarpa and Pinus albicaulus. Prosthecobacter, a hydrophilic bacterial genus, was more abundant on the green stripes, which had 6,524 bacterial sequences in comparison to the 1,183 sequences from the brown stripes. While further research can determine which functional traits are critical for these plants, knowledge of the current distribution of plant species and their functional traits can be used in predictive models of the responses of alpine plants to disappearing snowfields and glaciers. This research is important in conservation of rare arctic-alpine species on periglacial patterned ground.
APPLE Martha Elizabeth , RICKETTS Macy Kara , MARTIN Alice Caroline . Plant functional traits and microbes vary with position on striped periglacial patterned ground at Glacier National Park, Montana[J]. Journal of Geographical Sciences, 2019 , 29(7) : 1127 -1141 . DOI: 10.1007/s11442-019-1649-3
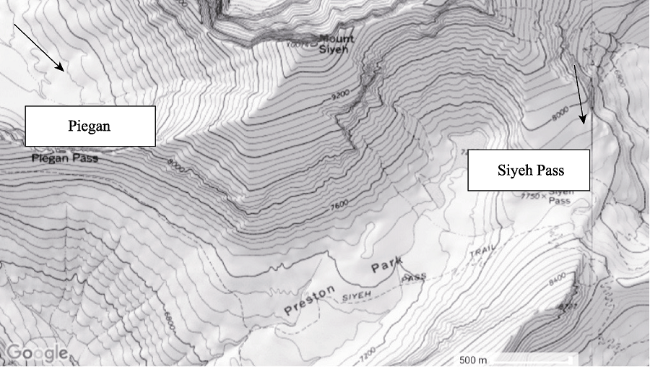
Figure 1 Google Earth Topographic Map of the Siyeh Pass and Piegan Pass snowfields at Glacier National Park, Montana |
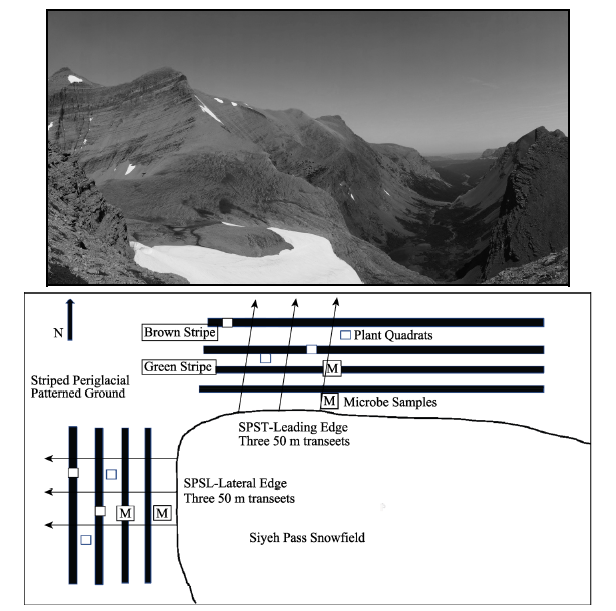
Figure 2 The Siyeh Pass Snowfield and adjacent periglacial patterned ground at Glacier National Park, Montana (above). Schematic diagram of sampling design (below). |
Table 1 The relative percent cover of qualitative plant functional traits on brown and green striped periglacial patterned ground at the Siyeh Pass Leading Edge (SPST), (n = 10 brown, n =10 green), and at the Siyeh Pass (SPSL) and Piegan Pass (PPSL) Lateral Edges (n =15 brown, n = 15 green). Shading of the higher value in a pair represents t-tests with significant differences of p ≤ 0.01 between brown and green stripes. VAM = Vesicular arbuscular mycorrhizae. |
| Average Relative Percent Cover +/- Standard Error | ||||||
|---|---|---|---|---|---|---|
| SPST | SPSL | PPSL | ||||
| Brown | Green | Brown | Green | Brown | Green | |
| Rare Species | 9.3 ± 2.0 | 0.19 ± 0.09 | 29.7 ± 0.3 | 0.25 ± 0.50 | 3.3 ± 1.2 | 0.07 ± 0.02 |
| Phanerophyte | 0 | 0 | 0 | 0 | 0 | 3.7 ± 0.4 |
| Hemicryptophyte | 86.9 ± 3.1 | 13.5 ± 1.8 | 91.7 ± 19.0 | 50.3 ± 3.9 | 72.0 ± 3.7 | 18.2 ± 1.7 |
| Chamaephyte | 15.7 ± 4.3 | 86.9 ± 1.5 | 0 | 48.2 ± 3.7 | 17.6 ± 3.7 | 67.1 ± 1.4 |
| Cushion | 22.4 ± 8.2 | 1.8 ± 0.3 | 30.7 ± 4.1 | 26.5 ± 2.6 | 7.4 ± 2.4 | 4.3 ± 8.9 |
| Mat | 29.2 ± 7.5 | 87.3 ± 0.8 | 0.36 ± 3.2 | 55.9 ± 3.2 | 45.2 ± 4.1 | 74.4 ± 1.5 |
| Rosette | 4.5 ± 2.5 | 1.4 ± 0.9 | 3.1 ± 3.7 | 0.5 ± 0.1 | 3.4 ± 0.9 | 0.8 ± 0.3 |
| Evergreen | 6.8 ± 2.4 | 69.6 ± 5.2 | 0 | 43.2 ± 3.5 | 12.1 ± 3.2 | 54.0 ± 3.0 |
| Simple Leaves | 75.4 ± 3.9 | 92.4 ± 1.2 | 61.6 ± 3.5 | 90.6 ± 1.4 | 97.5 ± 2.2 | 97.7 ± 3.5 |
| Lobed Leaves | 7.3 ± 1.6 | 0.7 ± 0.2 | 22.3 ± 3.1 | 0 | 2.3 ± 0.6 | 0.5 ± 0.1 |
| Clonality | 56.3 ± 4.3 | 92.6 ± 1.8 | 64.1 ± 1.6 | 96.3 ± 4.8 | 75.0 ± 2.9 | 81.6 ± 1.5 |
| Adventitious Roots | 49.5 ± 6.3 | 90.9 ± 1.6 | 0 | 48.2 ± 3.7 | 17.9 ± 3.8 | 69.6 ± 1.4 |
| Rhizomes | 32.1 ± 8.2 | 4.7 ± 0.3 | 88.6 ± 1.2 | 51.7 ± 3.7 | 76.8 ± 3.6 | 18.5 ± 1.4 |
| Taproots | 31.9 ± 7.3 | 2.1 ± 0.9 | 58.2 ± 4.0 | 31.2 ± 3.7 | 35.4 ± 2.5 | 10.7 ± 1.2 |
| Woody roots | 15.7 ± 4.3 | 86.9 ± 1.5 | 0 | 0 | 30.7 ± 8.3 | 2.6 ± 1.7 |
| Fibrous roots | 3.1 ± 1.7 | 0.1 ± 0.1 | 19.2 ± 1.4 | 3.1 ± 6.8 | 3.3 ± 0.1 | 0.4 ± 0.1 |
| Nitrogen fixers | 14.4 ± 3. | 74.7 ± 4.8 | 0.5 ± 0.1 | 85 ± 1.2 | 7.6 ± 2.8 | 26.5 ± 2.8 |
| All Mycorrhizae | 62.2 ± 5.9 | 93.1 ± 0.3 | 20.7 ± 2.2 | 78.7 ± 5.6 | 40.7 ± 6.3 | 80.0 ± 4.2 |
| VAM | 47.1 ± 5.6 | 7.4 ± 1.7 | 19.6 ± 2.0 | 37.3 ± 3.9 | 21.8 ± 3.5 | 29.4 ± 2.8 |
| Ectomycorrhizae | 20.0 ± 4.8 | 86.4 ± 2.1 | 12.3 ± 4.1 | 45.7 ± 4.4 | 16.4 ± 3.3 | 58.5 ± 4.4 |
Figure 3 The relative frequency of rare arctic-alpine species on brown and green stripes of periglacial patterned ground at Siyeh Pass and Piegan Pass at Glacier National Park |
Figure 4 The relative percent cover of rhizomatous and adventitiously-rooted species (mean +/- standard error) on brown and green stripes of periglacial patterned ground at Siyeh Pass and Piegan Pass at Glacier National Park |
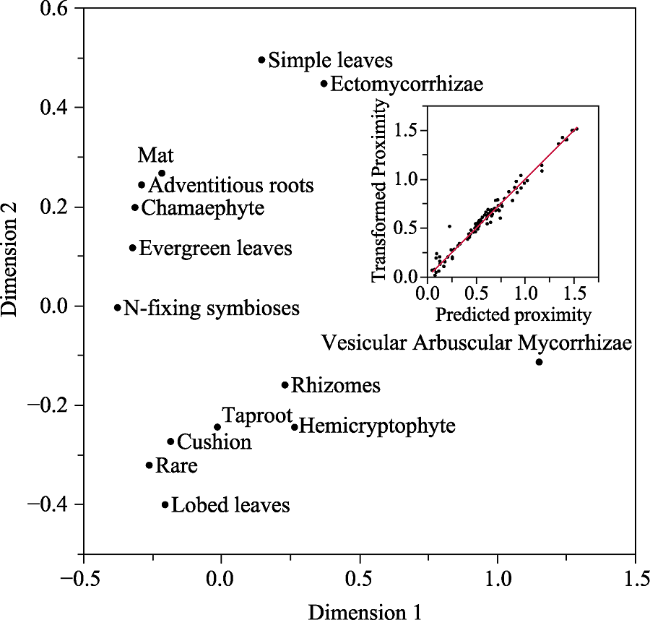
Figure 5 Multidimensional Scaling Plot of the relative percent cover of plant functional traits on striped periglacial patterned ground. The upper seven traits (Chamaephyte, Mat, Evergreen Leaves, Simple Leaves, Adventitious Roots, N-fixing symbioses, and Ectomycorrhizae) correspond with green stripes and the lower seven traits (Rare Species, Hemicryptophyte, Cushion, Lobed Leaves, Rhizomes, Taproots, and Vesicular Arbuscular Mycorrhizae) correspond with brown stripes. Details of fit are stress = 0.075, and R2 = 0.977 (in the inset Shepard Diagram). |
The authors have declared that no competing interests exist.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
Google Earth Version 7.1.8 3036, 2017. . 48°43’11”N, 113°37’47”W. 4.82 km, Siyeh Pass, Glacier National Park, Montana. March 1, 2018.
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|
| 54 |
|
| 55 |
|
| 56 |
|
| 57 |
|
| 58 |
|
| 59 |
|
| 60 |
|
| 61 |
|
| 62 |
|
| 63 |
|
| 64 |
|
| 65 |
|
| 66 |
|
| 67 |
|
| 68 |
|
| 69 |
|
| 70 |
|
| 71 |
|
| 72 |
|
| 73 |
|
| 74 |
|
| 75 |
|
| 76 |
|
| 77 |
|
| 78 |
|
| 79 |
|
| 80 |
|
| 81 |
|
| 82 |
|
| 83 |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}