
Journal of Geographical Sciences >
The response of spiders to less-focused non-crop habitats in the agricultural landscape along the lower reach of the Yellow River
*Corresponding author: Ding Shengyan, Professor, E-mail: syding@henu.edu.cn
Author: Hou Xiaoyun (1989-), PhD Candidate, specialized in ground arthropods research. E-mail: houxiaoyun526@126.com
Received date: 2018-05-10
Accepted date: 2019-01-22
Online published: 2019-07-25
Supported by
National Natural Science Foundation of China, No.41771202, No.41371195
Copyright
Non-crop habitats have been suggested to impact local biodiversity significantly in agricultural landscapes. However, there have been few studies of the effects of less-focused non-crop habitats (orchard, wetland, pit and ditch) on variation of spider abundance. In this study, spiders in 30 woodlands were captured using pitfall traps in Fengqiu County, China, and the effects of local and landscape variations at different scales (50 m, 100 m, 200 m, 350 m and 500 m) on spider abundance were analysed. The most important variation that influenced spider abundance at the 500 m scale was the less-focused non-crop habitat (LNH) cover, and 10% was an appropriate proportion of LNH cover to sustain high level of spider diversity in the investigated landscape. Non-metric multidimensional scaling analyses revealed that there were significant differences in the spider composition among the high, medium and low LNH coverage. Based on indicator species analysis, different spider species were associated with landscapes with different levels of LNH cover. Lycosidae, which accounted for 48% of the total specimens, preferred woodland habitats neighbouring areas with high LNH cover. Compared with woodland habitats, LNH provided more diverse food sources and habitat to sustain more spider species in the study area. Furthermore, linear elements composed of vegetation, such as pits and ditches, may prevent agricultural intensification by enhancing landscape connectivity and providing habitats for different spiders. Our findings may provide a theoretical basis for biodiversity conservation in agro-ecosystems and top-down control of pests.
HOU Xiaoyun , DING Shengyan , ZHAO Shuang , LIU Xiaobo . The response of spiders to less-focused non-crop habitats in the agricultural landscape along the lower reach of the Yellow River[J]. Journal of Geographical Sciences, 2019 , 29(7) : 1113 -1126 . DOI: 10.1007/s11442-019-1648-4
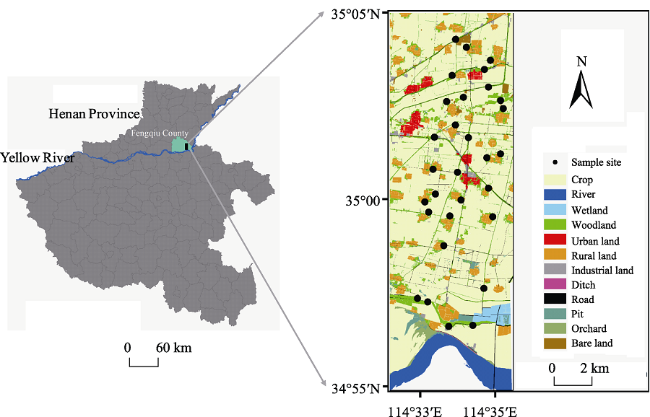
Figure 1 Distribution map of 30 sampling sites in the study area |
Table 1 Explanatory variables at the local (50 m) and landscape (100, 200, 350 and 500 m radii buffers) scales |
| Variables | Spatial scale (m) | Range (min-max)% | Mean ± SD |
|---|---|---|---|
| Crop cover | Local | 0-94 | 27.3±25.3 |
| Woodland cover | Local | 4-99 | 63±25.1 |
| Distance from road (m) | Local | 18.2-1269.5 | 434.1±397.2 |
| Crop cover | Landscape (100 m) | 8.1-91.2 | 44.1±20.3 |
| Woodland cover | Landscape (100 m) | 3-82 | 41.4±19.1 |
| LNH cover | Landscape (100 m) | 0-15.3 | 3.2±5.4 |
| Crop cover | Landscape (200 m) | 28-84.5 | 55.7±14.5 |
| Woodland cover | Landscape (200 m) | 5.2-56.7 | 26.2±12.1 |
| LNH cover | Landscape (200 m) | 0-30 | 3.4±6.5 |
| Crop cover | Landscape (350 m) | 35.5-81.5 | 60.4±11.4 |
| Woodland cover | Landscape (350 m) | 3-47.7 | 18.4±9.8 |
| LNH cover | Landscape (350 m) | 0-24.8 | 3.6±5.3 |
| Crop cover | Landscape (500 m) | 38.5-82.6 | 62.5±10.6 |
| Woodland cover | Landscape (500 m) | 3.9-42.8 | 15.5±8.6 |
| LNH cover | Landscape (500 m) | 0.2-19.8 | 4.1±6.1 |
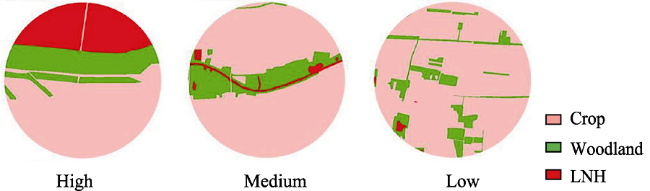
Figure 2 Three LNH cover groups at 500 m scale (each group has only one chosen typical site) |
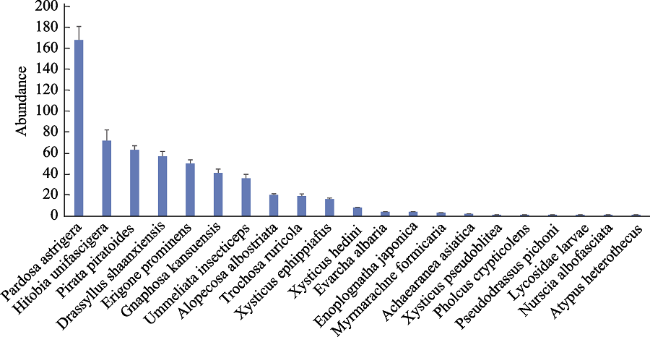
Figure 3 Abundance of spiders |
Table 2 Results of the information theoretic model selection and multi-model inference explaining spiders at the local (50 m) and landscape (100, 200, 350 and 500 m) scales |
| Model | Spatial scale | Adjusted R2 | Loglik | df | AICc | Wi | |
|---|---|---|---|---|---|---|---|
| Model 1 | 0.177 | -5.5798 | 8 | 32.2505 | |||
| Crop cover | Local | 0.29 | |||||
| Woodland cover | Local | 0.17 | |||||
| Distance from road (m) | Local | 0.18 | |||||
| Crop cover | Landscape (100 m) | 0.21 | |||||
| Woodland cover | Landscape (100 m) | 0.07 | |||||
| LNH cover | Landscape (100 m) | 0.06 | |||||
| Model 2 | 0.052 | -7.7082 | 8 | 36.5073 | |||
| Crop cover | Local | 0.34 | |||||
| Woodland cover | Local | 0.20 | |||||
| Distance from road (m) | Local | 0.21 | |||||
| Crop cover | Landscape (200 m) | 0.09 | |||||
| Woodland cover | Landscape (200 m) | 0.07 | |||||
| LNH cover | Landscape (200 m) | 0.08 | |||||
| Model 3 | 0.148 | -6.1067 | 8 | 33.3043 | |||
| Crop cover | Local | 0.30 | |||||
| Woodland cover | Local | 0.18 | |||||
| Distance from road (m) | Local | 0.19 | |||||
| Crop cover | Landscape (350 m) | 0.06 | |||||
| Woodland cover | Landscape (350 m) | 0.06 | |||||
| LNH cover | Landscape (350 m) | 0.21 | |||||
| Model 4 | 0.170 | -5.7152 | 8 | 32.5213 | |||
| Crop cover | Local | 0.24 | |||||
| Woodland cover | Local | 0.15 | |||||
| Distance from road (m) | Local | 0.15 | |||||
| Crop cover | Landscape (500 m) | 0.05 | |||||
| Woodland cover | Landscape (500 m) | 0.05 | |||||
| LNH cover | Landscape (500 m) | 0.36 |
Notes: Bolded print indicates the most powerful factors in each model. |
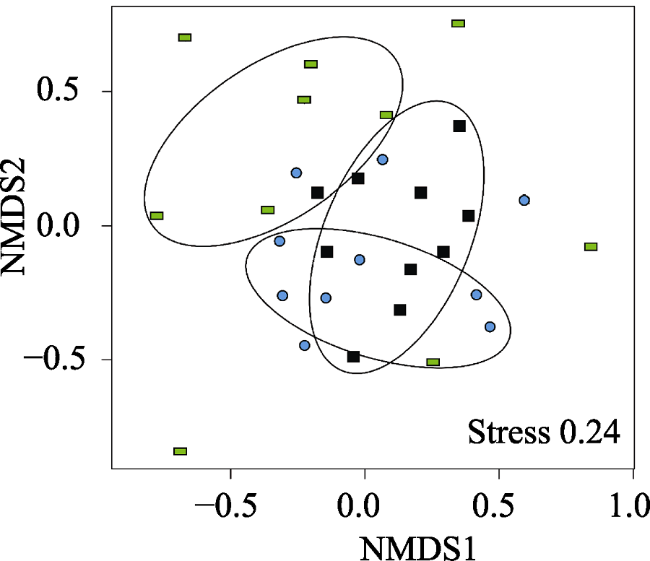
Figure 5 NMDS plot of three categories based on the spider composition at 500 m scale. Black symbols represent plots with a high LNH cover, blue symbols represent plots with a medium LNH cover, and green symbols represent plots with a low LNH cover. |
Table 3 Responses of spiders to LNH cover at 500 m scale by indicator species analysis |
| Taxa | Family | LNH coverage classes (500 m) and indicator values | ||
|---|---|---|---|---|
| Low | Medium | High | ||
| Pardosa astrigena | Lycosidae | 0.13 | 0.25 | 0.49 |
| Pirata piratoides | Lycosidae | 0.03 | 0.17 | 0.29 |
| Trochosa ruricola | Lycosidae | 0.01 | 0.08 | 0.48 |
| Alopecosa albostriata | Lycosidae | 0 | 0.02 | 0.44 |
| Lycosidae Larvae | Lycosidae | 0.10 | 0 | 0 |
| Gnaphosa kansuensis | Gnaphosidae | 0.10 | 0.13 | 0.34 |
| Drassyllus shaanxiensis | Gnaphosidae | 0.13 | 0.20 | 0.40 |
| Hitobia unifascigera | Gnaphosidae | 0.13 | 0.21 | 0.39 |
| Pseudodrassus pichoni | Gnaphosidae | 0 | 0 | 0.10 |
| Erigone prominens | Linyphiidae | 0.25 | 0.11 | 0.30 |
| Ummeliata insecticeps | Linyphiidae | 0.04 | 0.14 | 0.17 |
| Xysticus hedini | Thomisidae | 0 | 0.03 | 0.20 |
| Xysticus ephippiafus | Thomisidae | 0.11 | 0.05 | 0.11 |
| Xysticus pseudoblitea | Thomisidae | 0 | 0 | 0.10 |
| Evarcha albaria | Salticidae | 0 | 0.10 | 0.10 |
| Myrmarachne formicaria | Salticidae | 0 | 0.05 | 0.05 |
| Enoplognatha japonica | Theridiidae | 0.03 | 0.03 | 0.05 |
| Achaearanea asiatica | Theridiidae | 0 | 0.10 | 0 |
| Pholcus crypticolens | Pholcus | 0 | 0.10 | 0 |
| Nurscia albofasciata | Titanoecidae | 0 | 0 | 0.20 |
| Atypus heterothecus | Atypidae | 0 | 0 | 0.10 |
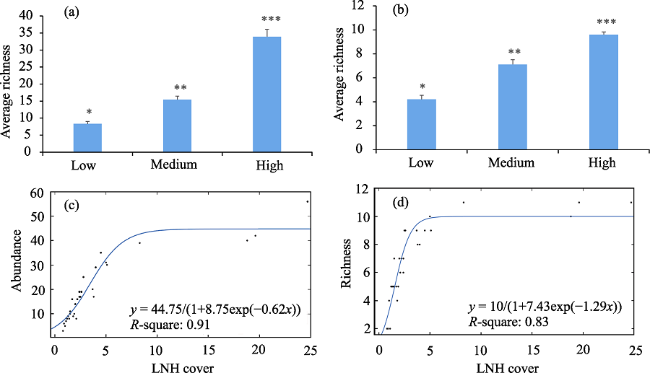
Figure 4 Abundance and richness of spiders in relation to LNH cover (500 m). The three LNH cover classes are based on the combination of orchard, wetland, pit and ditch areas at 500 m scale: 1) high LNH cover (High); 2) medium LNH cover (Medium); and 3) low LNH cover (Low). In the first row (a, b) the symbol (*) among LNH cover classes (High, Medium and Low) indicates significant differences in abundance and richness of spiders. The second row (c, d) shows the logistic trend-line of the abundance (or richness) and LNH cover. |
The authors have declared that no competing interests exist.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
MEA, 2005. Millennium Ecosystem Assessment. Washington: Island Press.
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|
| 54 |
|
| 55 |
R Development Core Team (RDCT), 2011. R: A language and environment for statistical computing. R foundation for statistical computing, Vienna, Austria. ISBN 3-900051-07-0, URL .
|
| 56 |
|
| 57 |
|
| 58 |
|
| 59 |
|
| 60 |
|
| 61 |
|
| 62 |
|
| 63 |
|
| 64 |
|
| 65 |
|
| 66 |
|
| 67 |
|
| 68 |
|
| 69 |
|
| 70 |
|
| 71 |
|
| 72 |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}