
Journal of Geographical Sciences >
The emerging cross-disciplinary studies of landscape ecology and biodiversity in China
Author: Shen Zehao, Professor, specialized in mountain landscape ecology and biogeography. E-mail: shzh@pku.edu.cn
Received date: 2018-05-10
Accepted date: 2019-01-22
Online published: 2019-07-25
Supported by
National Key Research and Development Program of China, No.2017YFC0505200
National Natural Science Foundation of China, No.41790425
Copyright
The spatiotemporal landscape heterogeneity implies multiple biodiversity mechanisms across scales, and the cross-disciplinary studies between landscape ecology and biodiversity are becoming a new research field in China. This paper briefly reviews the development of the field by comparing papers published in international journals and Chinese journals; then it investigates the differences in the trends and focuses between international and Chinese studies. We also introduce several study areas that have emerged over the last 10 years in this field, including metacommunity assembly, landscape genetics, biodiversity and ecosystem service relationship, and landscape planning for biodiversity conservation. The major advances emerging in this field in China over the past 5 years can be classified into six subject areas: 1) effects of urban landscape and urbanization on biodiversity; 2) altitudinal patterns of biodiversity in mountain landscapes; 3) effects of topographic heterogeneity on plant community assembly and species coexistence; 4) impacts of landscape patterns and processes on animal behaviors; 5) forest fires and spatiotemporal patterns of vegetation responses; and 6) landscape ecology applications in natural conservation planning and design. In an attempt to promote cross-field studies between geography and ecology, this special issue collected 10 research articles, involving multiple landscape types and biological assemblages, in order to explore the interaction between landscape features and biodiversity. We anticipate that the future development of this active front will be primarily driven by the application of novel information techniques and the realistic demands of sustainability issues, in addition to answering scientific questions cross scales.
Key words: landscape ecology; biodiversity; cross-disciplinary study; frontiers; progresses
SHEN Zehao , LI Yiying , YANG Kang , CHEN Lifang . The emerging cross-disciplinary studies of landscape ecology and biodiversity in China[J]. Journal of Geographical Sciences, 2019 , 29(7) : 1063 -1080 . DOI: 10.1007/s11442-019-1645-7
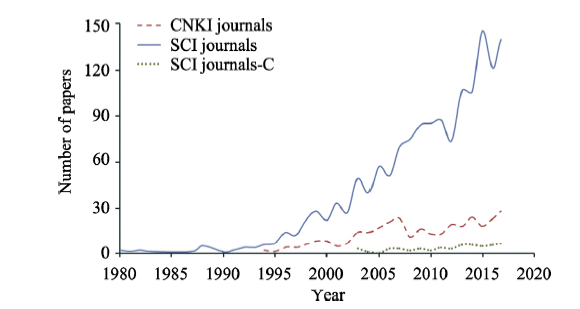
Figure 1 Temporal change in publications on the “landscape + biodiversity” topic. SCI journal-C: papers in SCI journals contributed by authors from Chinese institutions |
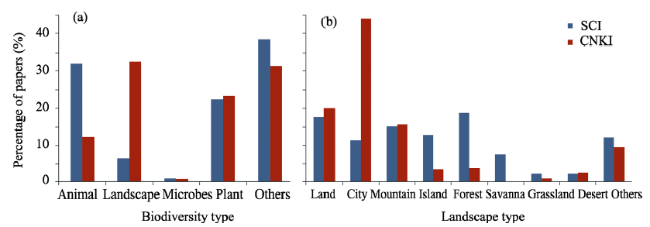
Table 1 The frequency distributions of “Landscape + biodiversity” papers published in SCI journals and CNKI journals, with respect to the corresponding biodiversity and landscape categories |
| Object | Category | SCI journal | CNKI journal | ||
|---|---|---|---|---|---|
| Number of papers | % | Number of papers | % | ||
| Landscape context | Land | 276 | 17.5 | 56 | 20.1 |
| City | 180 | 11.4 | 122 | 44.0 | |
| Mountain | 240 | 15.2 | 44 | 15.8 | |
| Island | 203 | 12.9 | 9 | 3.3 | |
| Forest | 298 | 18.9 | 11 | 3.8 | |
| Savanna | 121 | 7.7 | 1 | 0.3 | |
| Grassland | 33 | 2.1 | 3 | 1.0 | |
| Desert | 33 | 2.1 | 7 | 2.4 | |
| Others | 191 | 12.1 | 27 | 9.6 | |
| Biodiversity type | Animal | 502 | 31.8 | 34 | 12.2 |
| Landscape | 102 | 6.5 | 90 | 32.4 | |
| Microbes | 14 | 0.9 | 2 | 0.7 | |
| Plant | 353 | 22.4 | 65 | 23.4 | |
| Others | 606 | 38.4 | 87 | 31.3 | |
| Total | 1577 | 100.0 | 278 | 100.0 | |
Figure 2 Comparisons of the percentage composition of papers published in SCI and CNKI journals that addressed (a) biodiversity type and (b) landscape type subject areas |
Table A1 The interannual change of numbers of papers published in CNKI journals, SCI journals, as well as those SCI papers contributed by Chinese institutes (SCI journal-C) |
| Publishing year | SCI journal | CNKI journal | SCI journal-C |
|---|---|---|---|
| 1980 | 2 | ||
| 1981 | 1 | ||
| 1982 | 2 | ||
| 1983 | 1 | ||
| 1987 | 1 | ||
| 1988 | 5 | ||
| 1990 | 1 | ||
| 1991 | 2 | ||
| 1992 | 4 | ||
| 1993 | 4 | ||
| 1994 | 6 | 2 | |
| 1995 | 7 | 1 | |
| 1996 | 14 | 4 | |
| 1997 | 12 | 4 | |
| 1998 | 22 | 6 | |
| 1999 | 28 | 8 | |
| 2000 | 22 | 8 | |
| 2001 | 33 | 5 | |
| 2002 | 27 | 7 | |
| 2003 | 49 | 14 | 3 |
| 2004 | 40 | 14 | 1 |
| 2005 | 57 | 17 | 0 |
| 2006 | 51 | 21 | 3 |
| 2007 | 70 | 23 | 3 |
| 2008 | 75 | 11 | 2 |
| 2009 | 84 | 16 | 3 |
| 2010 | 85 | 13 | 2 |
| 2011 | 87 | 13 | 4 |
| 2012 | 74 | 19 | 3 |
| 2013 | 107 | 18 | 6 |
| 2014 | 106 | 24 | 6 |
| 2015 | 145 | 18 | 5 |
| 2016 | 121 | 23 | 6 |
| 2017 | 138 | 28 | 7 |
The authors have declared that no competing interests exist.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|
| 54 |
|
| 55 |
|
| 56 |
|
| 57 |
|
| 58 |
|
| 59 |
|
| 60 |
|
| 61 |
|
| 62 |
|
| 63 |
|
| 64 |
|
| 65 |
|
| 66 |
|
| 67 |
|
| 68 |
|
| 69 |
|
| 70 |
|
| 71 |
|
| 72 |
|
| 73 |
|
| 74 |
|
| 75 |
|
| 76 |
|
| 77 |
|
| 78 |
|
| 79 |
|
| 80 |
|
| 81 |
|
| 82 |
|
| 83 |
|
| 84 |
|
| 85 |
|
| 86 |
|
| 87 |
|
| 88 |
|
| 89 |
|
| 90 |
|
| 91 |
|
| 92 |
|
| 93 |
|
| 94 |
|
| 95 |
|
| 96 |
|
| 97 |
|
| 98 |
|
| 99 |
|
| 100 |
|
| 101 |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}