
Journal of Geographical Sciences >
Potential priority areas and protection network for Yunnan snub-nosed monkey (Rhinopithecus bieti) in Southwest China
*Corresponding author: Liu Guohua, E-mail: ghliu@rcees.ac.cn
Author: Su Xukun, PhD, specialized in biodiversity conservation and landscape ecology. E-mail: xksu@rcees.ac.cn
Received date: 2018-05-10
Accepted date: 2019-01-22
Online published: 2019-07-25
Supported by
National Key Research and Development Program of China, No.2016YFC0502102
Copyright
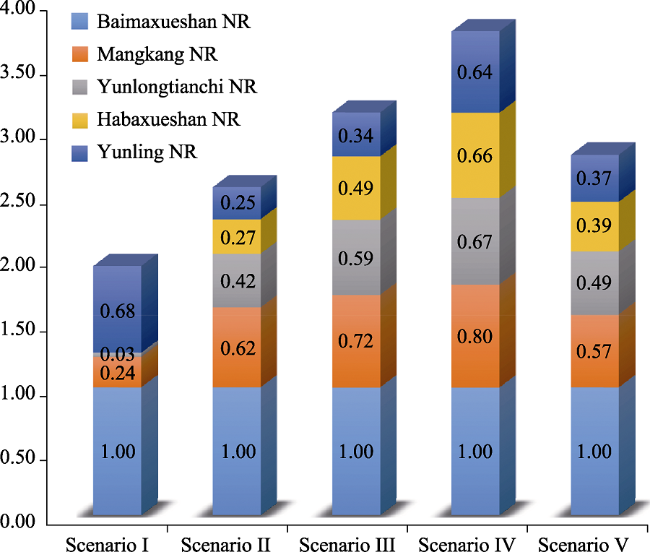
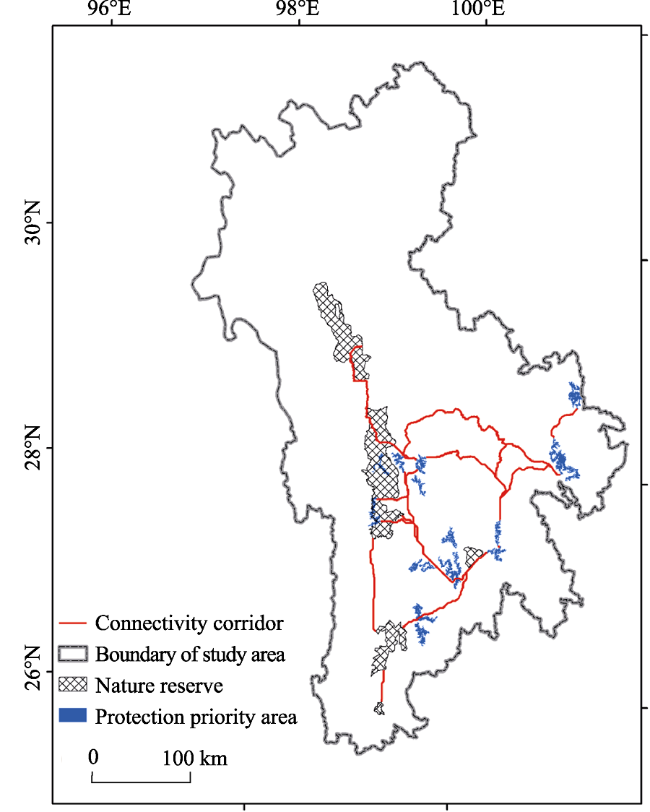
In Southwest China, five Nature Reserves (NRs) (Mangkang, Baimaxueshan, Yunling, Habaxueshan, and Yunlongtianchi) play a key role in protecting the endemic and endangered Yunnan snub-nosed monkey (YSM) (Rhinopithecus bieti). However, increasing human activities threaten its habitats and corridors. We used a GIS-based Niche Model to delineate potential core habitats (PCHs) of the YSMs and a Linkage Mapper corridor simulation tool to restore potential connectivity corridors (PCCs), and defined five scenarios. A normalized importance value index (NIVI) was established to identify the protection priority areas (PPAs) for the YSMs for five scenarios. The results indicated that locations of the habitats and corridors were different in the five scenarios, thereby influencing the distribution of the PPAs and protection network of the YSMs. The NIVI value of Baimaxueshan nature reserve was 1 in the five scenarios, which implied the maximum importance. There were only 7 PCHs and 16 PCCs (with the longest average length of 223.13 km) which were mainly located around 5 NRs in scenario III. The protection network of the YSMs was composed of 16 PCHs, 18 PCCs, and 5 NRs. Under each scenario, most of the PCHs and the PCCs were located in the south of the study area. The five NRs only covered 2 PPAs of the YSMs. We suggest that the southern part of the study area needs to be strictly protected and human activities should be limited. The area of the five NRs should be expanded to maximize protection of the YSMs in the future.
SU Xukun , HAN Wangya , LIU Guohua . Potential priority areas and protection network for Yunnan snub-nosed monkey (Rhinopithecus bieti) in Southwest China[J]. Journal of Geographical Sciences, 2019 , 29(7) : 1211 -1227 . DOI: 10.1007/s11442-019-1654-6
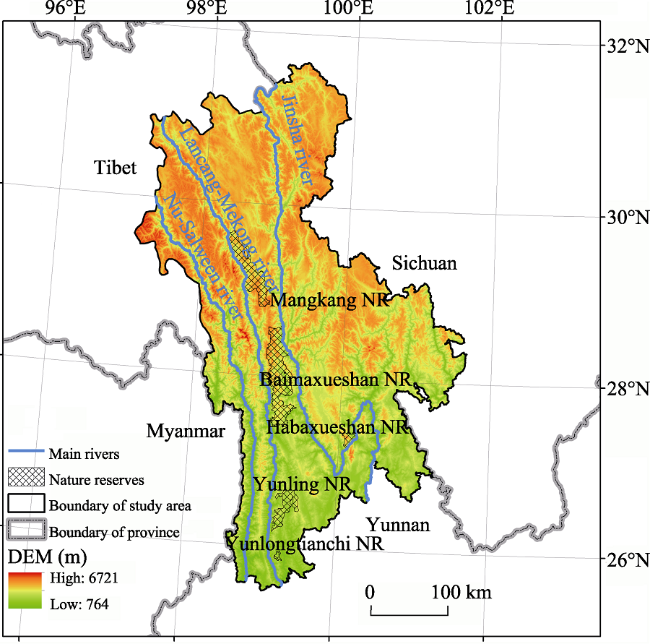
Figure 1 Location of the study area |
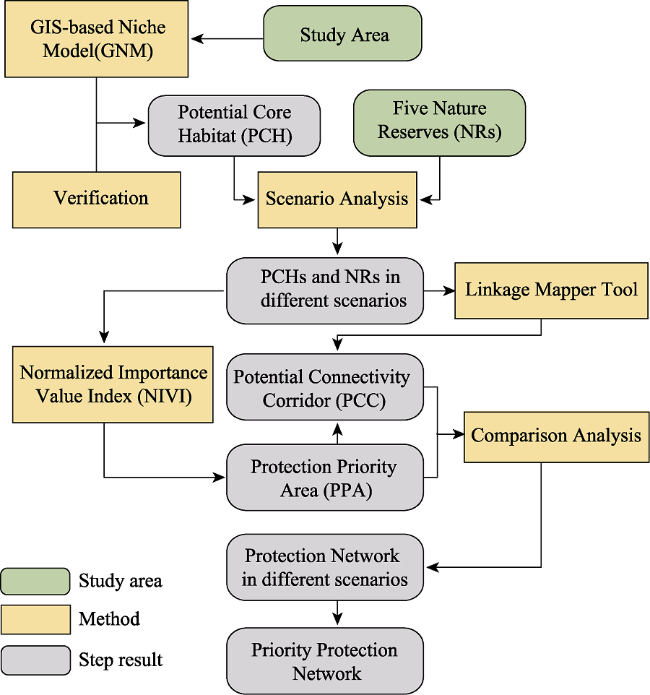
Figure 2 Technical flowchart |
Table 1 Weights of each variable |
| Vegetation types | ||||||
|---|---|---|---|---|---|---|
| Name | Value | Name | Value | |||
| Tsuga chinensis pritz and Betula alnoides | 100 | Cotinus nana | 50 | |||
| Tsuga dumosa (D. Don) Eichler | 100 | Sophora davidii | 50 | |||
| Betula platyphylla Suk | 75 | Imperata cylindrica | 50 | |||
| Abies delavayi Franch | 75 | Bothriochloa ischaemum (L.) Keng | 50 | |||
| Betula albo-sinensis Burk. var. septentrionalis Schneid | 75 | Rhododendron telmateium | 50 | |||
| Platycladus orientalis | 75 | Arundinella setosa, Arundinella anomala | 50 | |||
| Quercus aquifolioides | 75 | Rhododendron flavidum Franch | 50 | |||
| bies forrestii | 75 | Salix cupularis | 50 | |||
| Picea balfouriana | 75 | Themeda triandra Forsk. Var. Japonica (Willd. )Makino and Miscanthus | 50 | |||
| Neosino calamus affinis | 75 | Caragana jubata (Pall.) Poir. | 50 | |||
| Larix potaninii var. macrocarpa | 75 | Rosa sericea Lindl. | 50 | |||
| Sabina tibetica Kom | 75 | Rhododendron heliolepis Franch. | 50 | |||
| Lithocarpus variolosus (Fr.) Chun | 75 | Rhododendron delavayi Franch. | 50 | |||
| Castanopsis delavayi Franch | 75 | Rhododendron fastigiatum Franch. | 50 | |||
| Pinus densata | 75 | Vaccinium bracteatum Thunb. | 50 | |||
| Quercus pseudosemecarpifolia A. Camus | 75 | Heteropogon contortus | 50 | |||
| Betula albo-sinensis Burk | 75 | Malus baccata (L.) Borkh, Prunus padus L. | 50 | |||
| Pinus armandii Franch | 75 | Rhododendron adenogynum Diels | 50 | |||
| Quercus pannosa Hand.-Mazz. | 75 | Sabina pingiivar.wilsonii and Sabina squamata | 50 | |||
| Fargesia spathacea Franch | 75 | Potentilla parvifolia | 50 | |||
| Picea likiangensis | 75 | Rhododendron nivaleHook. f. | 50 | |||
| Abies squamata Mast | 75 | Rhododendron racemosum Franch | 50 | |||
| Pinus massoniana Lamb | 75 | Salix sclerophylla Anderss | 50 | |||
| Quercus guyavaefolia Levl | 75 | Phyllanthus emblica L. | 50 | |||
| Abies faxoniana Rehd | 75 | Rhododendron morii Hayata | 50 | |||
| Pobulus davidiana | 75 | Sibiraea angustata (Rehd.) Hand.-Mazz. | 50 | |||
| Quercus variabilis Bl | 75 | Kobresia littledalei C. B. Clarke | 25 | |||
| Larix chinensis | 75 | Carex liparocarpos Gaudin | 25 | |||
| Abies spectabilis (D. Don) Spach | 75 | Anaphalis flavescens Hand.-Mazz. | 25 | |||
| Castanopsisindica (Roxb.) Miq, Castanopsis clarkei | 75 | Sanguisorba officinalisL. and Artemisia tanacetifolia Linn. | 25 | |||
| Piceabrachytylavar.omplanata | 75 | B. sylvaticum (Huds) Beauv | 25 | |||
| Castanopsis concolor Rehd. et Wils. | 75 | Blysmus sinocompressus Tang et Wang | 25 | |||
| Pinus yunnanensis | 75 | Cyperaceae | 25 | |||
| Picea asperata Mast. | 75 | Kobresia humilis and Polygonum macrophyllum D. Don | 25 | |||
| Abies georgei Orr | 75 | Festuca ovina L. and Deyeuxia arundinacea | 25 | |||
| Pinus palustris Mill. | 75 | Polygonum macrophyllum D. Don and Polygonum viviparum L | 25 | |||
| Picea purpurea Mast | 75 | Poa annua L. | 25 | |||
| Quercus monimotricha | 50 | Non-vegetation | 0 | |||
| Land cover | ||||||
| Name | Value | Name | Value | |||
| Forest | 100 | Wetland | 0 | |||
| Shrub | 75 | Lake | 0 | |||
| Meadow | 25 | River | 0 | |||
| Steppe | 25 | Farmland | 0 | |||
| Vegetation types | ||||||
| Forest wetland | 50 | Settlement | 0 | |||
| Sparse bushes | 50 | Road | 0 | |||
| Shrub wetland | 0 | |||||
| Aspect | ||||||
| Class | Value | Class | Value | |||
| 0 | 0 | 136°-225° | 100 | |||
| 1-45° | 25 | 226°-270° | 75 | |||
| 46°-90° | 50 | 271°-315° | 50 | |||
| 91°-135° | 75 | |||||
| Slope | ||||||
| Class | Value | Class | Value | |||
| 0° | 0 | 20°-40° | 100 | |||
| 0°-90° | 25 | Above 40° | 25 | |||
| 15°-20° | 50 | |||||
| Water | ||||||
| Distance | Value | Distance | Value | |||
| Water source | 0 | 3.5 km away from water source | 50 | |||
| 1 km away from water source | 100 | More than 3.5 km away from water source | 25 | |||
| Elevation | ||||||
| Class | Value | Class | Value | |||
| 764-2600 m, 4501-4700 m | 25 | 2801-3600 m | 100 | |||
| 2601-2700 m, 4301-4500 m | 50 | Above 4701 m | 0 | |||
| 2701-2800 m, 3601-4300 m | 75 | |||||
| Settlement | ||||||
| Class | Value | Class | Value | |||
| 1 km away from settlement | 0 | More than 3 km away from settlement | 100 | |||
| 3 km away from settlement | 50 | |||||
| Road | ||||||
| Distance | Value | Distance | Value | |||
| 50 m away from road | 0 | More than 100 m away from road | 100 | |||
| 100 m away from road | 50 | |||||
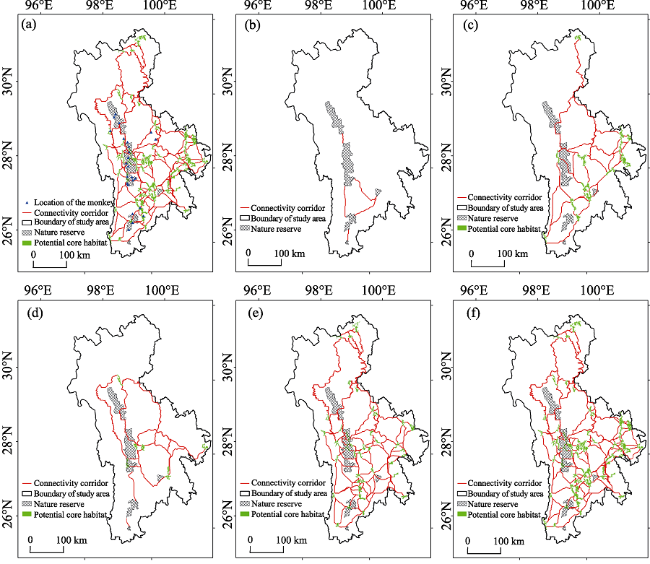
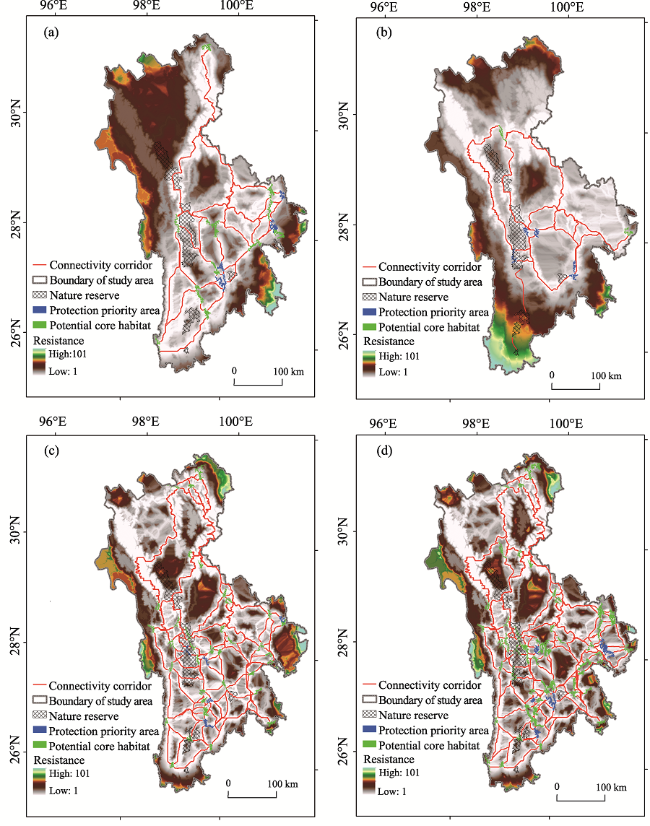
Figure 3 Existing location of the YSMs in the study area (a); location of the PCCs in scenario I (b); distribution of the PCHs and PCCs in scenario II (c); distribution of the PCHs and PCCs in scenario III (d); distribution of the PCHs and PCCs in scenario IV (e); distribution of the PCHs and PCCs in scenario V (f) |
Table 2 Total number, total area and average area of the PCHs and NRs in different scenarios |
| Total number | Total area (km2) | Average area (km2) | |
|---|---|---|---|
| Scenario I | 5 | 5679 | 1135.80 |
| Scenario II | 16 | 2121 | 132.56 |
| Scenario III | 7 | 635 | 90.71 |
| Scenario IV | 40 | 2396 | 59.90 |
| Scenario V | 63 | 5152 | 81.78 |
Table 3 Total number, total distance and average length of the PCCs in different scenarios |
| Total number | Total distance (km) | Average length (km) | |
|---|---|---|---|
| Scenario I | 5 | 391.68 | 78.34 |
| Scenario II | 31 | 4329.05 | 139.65 |
| Scenario III | 16 | 3570.07 | 223.13 |
| Scenario IV | 122 | 12126.41 | 99.40 |
| Scenario V | 182 | 11842.43 | 65.07 |
Figure 4 The NIVI value changes of the five NRs in each scenario |
Figure 5 Distribution of the PPAs and the PCCs in scenario II (a), scenario III (b); scenario IV (c), and scenario V (d) |
Figure 6 Protection network of the YSMs |
The authors have declared that no competing interests exist.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}