
Journal of Geographical Sciences >
Spatial patterns and environmental factors influencing leaf carbon content in the forests and shrublands of China
Author: Zhao Hang (1991-), PhD, specialized in plant carbon and nitrogen storage. E-mail: zhh1108@163.com
Received date: 2017-05-31
Accepted date: 2017-10-17
Online published: 2018-06-20
Supported by
National Key R&D Program of China, No.2016YFA0600103, No.2017YFA0604803
Youth Innovation Research Team Project, No.LENOM2016Q0005
Copyright
Leaf carbon content (LCC) is widely used as an important parameter in estimating ecosystem carbon (C) storage, as well as for investigating the adaptation strategies of vegetation to their environment at a large scale. In this study, we used a dataset collected from forests (5119 plots) and shrublands (2564 plots) in China, 2011-2015. The plots were sampled following a consistent protocol, and we used the data to explore the spatial patterns of LCC at three scales: plot scale, eco-region scale (n = 24), and eco-region scale (n = 8). The average LCC of forests and shrublands combined was 45.3%, with the LCC of forests (45.5%) being slightly higher than that of shrublands (44.9%). Forest LCC ranged from 40.2% to 51.2% throughout the 24 eco-regions, while that of shrublands ranged from 35% to 50.1%. Forest LCC decreased with increasing latitude and longitude, whereas shrubland LCC decreased with increasing latitude, but increased with increasing longitude. The LCC increased, to some extent, with increasing temperature and precipitation. These results demonstrate the spatial patterns of LCC in the forests and shrublands at different scales based on field-measured data, providing a reference (or standard) for estimating carbon storage in vegetation at a regional scale.
Key words: carbon storage; eco-regions; foliar carbon; shrubs; stoichiometry; China
ZHAO Hang , XU Li , WANG Qiufeng , TIAN Jing , TANG Xuli , TANG Zhiyao , XIE Zongqiang , HE Nianpeng , YU Guirui . Spatial patterns and environmental factors influencing leaf carbon content in the forests and shrublands of China[J]. Journal of Geographical Sciences, 2018 , 28(6) : 791 -801 . DOI: 10.1007/s11442-018-1505-x
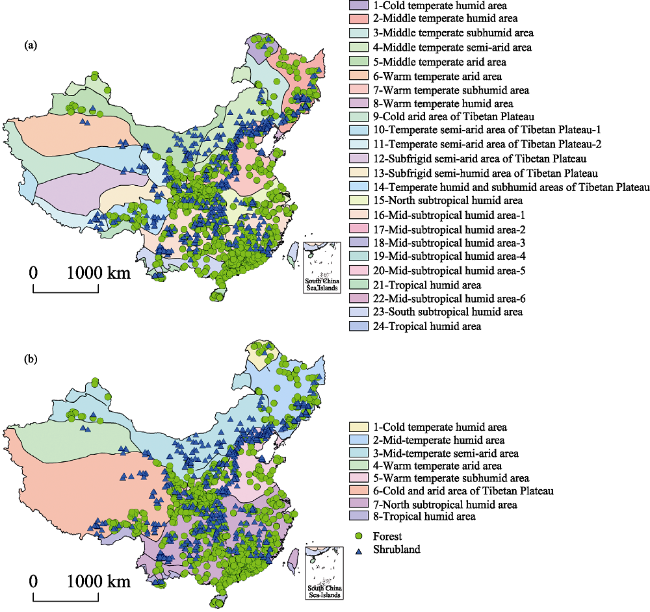
Figure 1 Sampling sites of forests and shrublands in China divided into 24 eco-regions (a) and 8 eco-regions (b) |
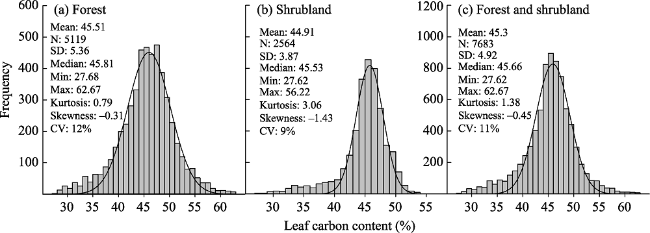
Figure 2 Frequency distribution of leaf carbon content (LCC, %) in the forests and shrublands of China (N, sample number; SD, standard deviation; Min, minimum value; Max, maximum value; CV, coefficient of variation) |
Table 1 Changes in leaf carbon content (LCC, %) of forests and shrublands in the 24 eco-regions of China |
| Region | LCC of forests | LCC of shrublands | LCC of forests and shrublands | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Mean (%) | N ‡ | SD | Mean (%) | N | SD | Mean (%) | N | SD | |
| 1† | 42.73Ag§ | 29 | 7.44 | 46.10Abcde | 6 | 3.18 | 43.31g | 35 | 6.98 |
| 2 | 40.18Ah | 541 | 6.96 | 44.13Be | 299 | 2.35 | 41.59h | 840 | 6.06 |
| 3 | 41.56Ag | 51 | 5.95 | 45.8Bbcd | 19 | 1.29 | 42.71g | 70 | 5.45 |
| 4 | 46.84Ade | 197 | 4.85 | 45.19Bcd | 245 | 2.32 | 45.93d | 442 | 3.76 |
| 5 | 45.71Aef | 44 | 4.38 | 39.84Bf | 230 | 5.34 | 40.78i | 274 | 5.62 |
| 6 | - | - | - | 36.96g | 24 | 3.87 | 36.96j | 24 | 3.87 |
| 7 | 45.17Af | 645 | 4.54 | 45.47Ac | 458 | 2.00 | 45.30ef | 1103 | 3.70 |
| 8 | 49.18bcd | 27 | 2.22 | - | - | - | 49.18ab | 27 | 2.22 |
| 9 | - | - | - | - | - | - | - | - | - |
| 10 | - | - | - | 34.99h | 54 | 4.38 | 34.99j | 54 | 4.38 |
| 11 | 49.02Abc | 45 | 4.03 | 46.14Bb | 148 | 4.08 | 46.81cd | 193 | 4.24 |
| 12 | - | - | 43.85e | 3 | 0.71 | 43.85fghi | 3 | 0.71 | |
| 13 | - | - | - | 45.77bc | 64 | 3.15 | 45.77def | 64 | 3.15 |
| 14 | 48.21Ac | 266 | 3.73 | 48.06Aa | 81 | 3.15 | 48.17b | 347 | 3.60 |
| 15 | 45.52Aef | 651 | 3.46 | 44.78Bd | 118 | 1.96 | 45.41e | 769 | 3.28 |
| 16 | 46.02Ae | 199 | 3.40 | 45.47Acd | 96 | 1.81 | 45.84de | 295 | 2.98 |
| 17 | 42.62Ag | 265 | 5.14 | 45.66Bbc | 157 | 2.68 | 43.75g | 422 | 4.62 |
| 18 | 46.73Ade | 297 | 4.14 | 45.92Bbc | 149 | 2.32 | 46.46d | 446 | 3.65 |
| 19 | 44.79Af | 661 | 4.10 | 47.01Bab | 46 | 2.28 | 44.94f | 707 | 4.05 |
| 20 | 51.24Aa | 76 | 3.44 | 48.03Ba | 257 | 1.92 | 48.77b | 333 | 2.71 |
| 21 | 49.26Abc | 44 | 2.93 | 50.12Aa | 3 | 0.62 | 49.32ab | 47 | 2.84 |
| 22 | 49.69Ab | 342 | 5.78 | 45.52Bbcde | 17 | 1.48 | 49.49a | 359 | 5.72 |
| 23 | 47.38Ad | 478 | 4.58 | 46.48Ab | 40 | 2.85 | 47.31c | 518 | 4.47 |
| 24 | 45.28Aef | 261 | 4.36 | 44.67Ade | 50 | 1.85 | 45.18ef | 311 | 4.07 |
† Eco-region number corresponding with Figure 1a ‡ N = number of sampling sites; SD = standard deviation § Different uppercase letters indicate significant differences between forests and shrublands within specific rows at P < 0.05; Different lowercase letters indicate significant differences among the 24 eco-regions within each column at P < 0.05 |
Table 2 Leaf carbon content (LCC, %) of forests and shrublands in the eight larger eco-regions of China |
| Area | LCC of forests | LCC of shrublands | LCC of forests and shrublands | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Mean (%) | N ‡ | SD | Mean (%) | N | SD | Mean (%) | N | SD | |
| 1† | 42.73Ae§ | 29 | 7.44 | 46.10Aabcd | 6 | 3.18 | 43.31d | 35 | 6.98 |
| 2 | 40.30Af | 592 | 6.89 | 44.23Bd | 318 | 2.33 | 41.67e | 910 | 6.02 |
| 3 | 46.63Ab | 241 | 4.78 | 42.60Be | 475 | 4.87 | 43.96d | 716 | 5.20 |
| 4 | - | - | - | 36.96f | 24 | 3.87 | 36.96f | 24 | 3.87 |
| 5 | 45.33Ad | 672 | 4.53 | 45.47Ab | 458 | 2.00 | 45.39c | 1130 | 3.72 |
| 6 | 48.32Aa | 311 | 3.78 | 44.78Bc | 350 | 5.68 | 46.44a | 661 | 5.19 |
| 7 | 45.95Ac | 2491 | 4.77 | 46.36Ba | 840 | 2.47 | 46.05b | 3331 | 4.31 |
| 8 | 46.65Ab | 743 | 4.61 | 46.70Aa | 133 | 3.18 | 46.66a | 876 | 4.42 |
† Eco-region number corresponding with Figure 1b ‡ N = number of sampling sites; SD = standard deviation § Different uppercase letters indicate significant differences between forests and shrublands within specific rows at P < 0.05; Different lowercase letters indicate significant differences among the 8 larger eco-regions within each column at P < 0.05. |
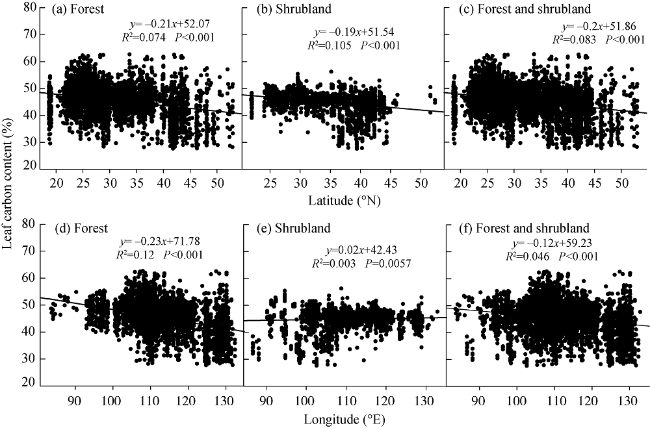
Figure 3 Changes in leaf carbon content (LCC, %) with latitude and longitude in the forests and shrublands of China at the plot scale |
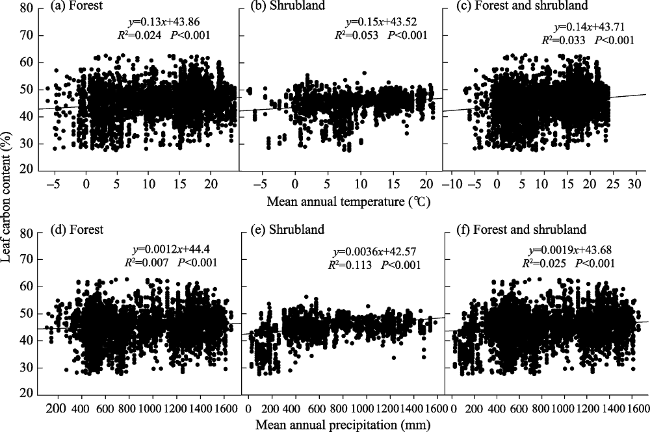
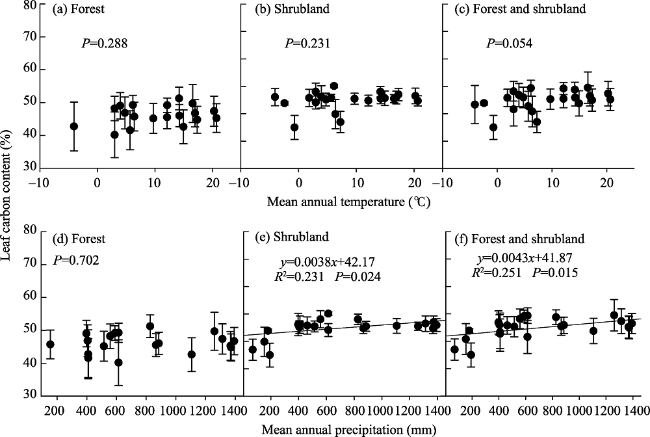
Figure 4 Changes in leaf carbon content (LCC, %) in the forests and shrublands of China with climate at the plot scale |
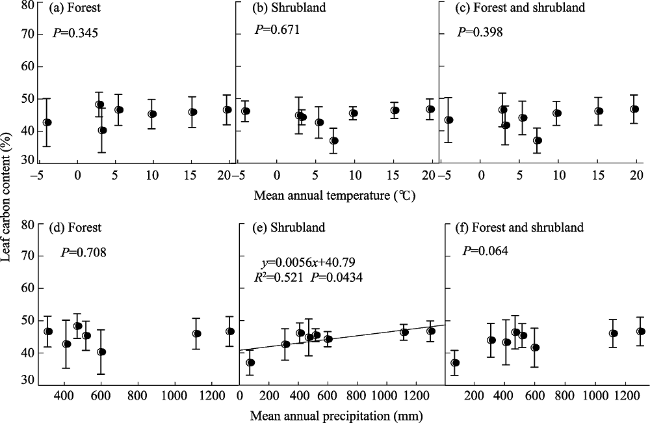
Figure 5 Changes in leaf carbon content (LCC %) in the forests and shrublands of China with climate at the scale of the 24 eco-regions |
Figure 6 Changes in leaf carbon content (LCC, %) in the forests and shrublands of China with climate at the scale of the eight eco-regions |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}