
Journal of Geographical Sciences >
Dynamic changes of habitats in China’s typical national nature reserves on spatial and temporal scales
Author: Zhu Ping, Master Student, specialized in remote sensing of ecology. E-mail: zhup.15s@igsnrr.ac.cn
Received date: 2017-10-27
Accepted date: 2017-12-30
Online published: 2018-06-20
Supported by
The National Key Research and Development Program, No.2017YFC0506404
The Key Programs for Frontier Science of the Chinese Academy of Sciences, No.QYZDB-SSW-DQC005
The National Science & Technology Pillar Program, No.2013BAC03B00
Copyright
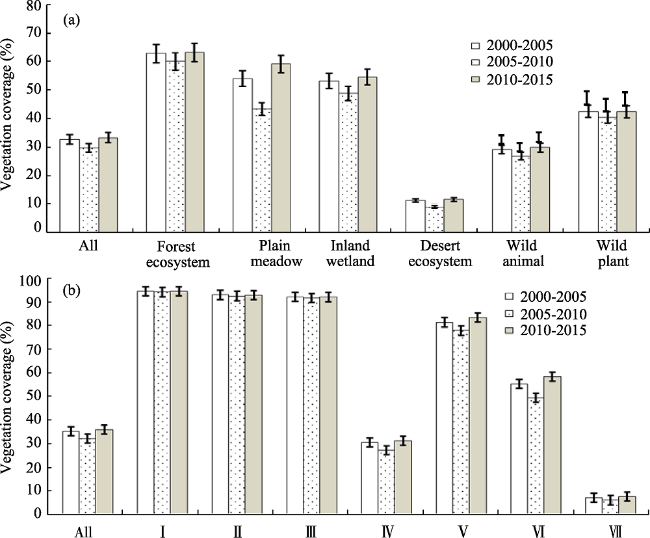
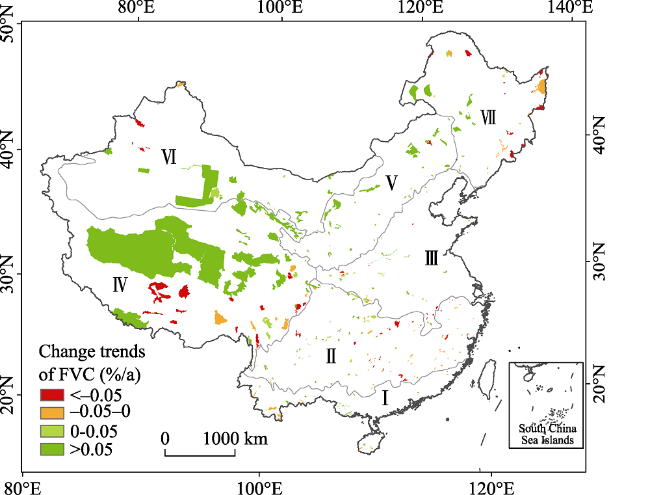
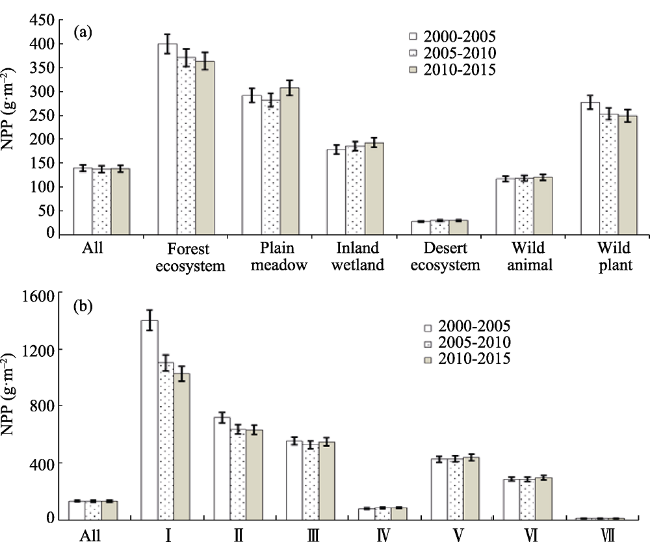
Until 2015, China had established 2740 nature reserves with a total area of 1.47 million km2, covering 14.8% of China’s terrestrial land surface. Based on remote sensing inversion, ecological model simulation and spatial analysis methods, we analyzed the spatial and temporal variations of fractional vegetation coverage (FVC), net primary production (NPP), and human disturbance (HD) in habitats of typical national nature reserves (NNRs) during the first 15 years of the 21st century from 2000 to 2015. And then the three indicators were compared between different NNR types and varied climate zones. The results showed that (1) the average 5-year FVC of NNRs increased from 36.3% to 37.1%, and it improved in all types of NNRs to some extent. The annual average FVC increased by 0.11%, 0.84%, 0.21%, 0.09%, 0.11% and 0.08% in NNRs of forest ecosystem, plain meadow, inland wetland, desert ecosystem, wild animal and wild plant, respectively. (2) The NPP annually increased by 2.06 g·m-2, 1.23 g·m-2, 0.28 g·m-2 and 0.4 g·m-2 in NNRs of plain meadow, inland wetland, desert ecosystem and wild animal, respectively. However, it decreased by 3.45 g·m-2 and 2.35 g·m-2 in NNRs of forest ecosystem and wild plant respectively. (3) In the past 15 years, besides the slight decreases in the NNRs located at the Qinghai-Tibet Plateau and the south subtropical zone, HD enhanced in most of NNRs, especially HD in the warm temperate humid zone increased from 4.7% to 5.35%.
ZHU Ping , HUANG Lin , XIAO Tong , WANG Junbang . Dynamic changes of habitats in China’s typical national nature reserves on spatial and temporal scales[J]. Journal of Geographical Sciences, 2018 , 28(6) : 778 -790 . DOI: 10.1007/s11442-018-1504-y
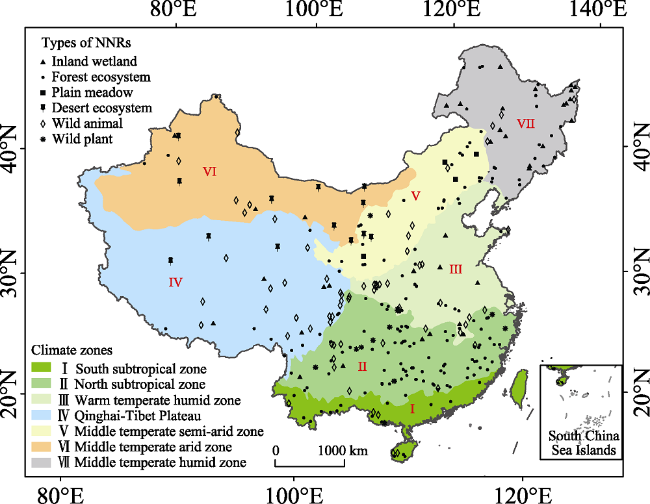
Figure 1 Spatial distribution of 299 typical NNRs in China |
Table 1 Conversion factor of different land use and land cover types |
| Land use and land cover | Cropland | Artificial forest | Reservoir | Built-up areas | Others |
|---|---|---|---|---|---|
| Conversion factor | 0.2 | 0.133 | 0.6 | 1 | 0 |
Figure 2 The average FVC in typical NNRs (a) with different types and (b) located in varied climate zones |
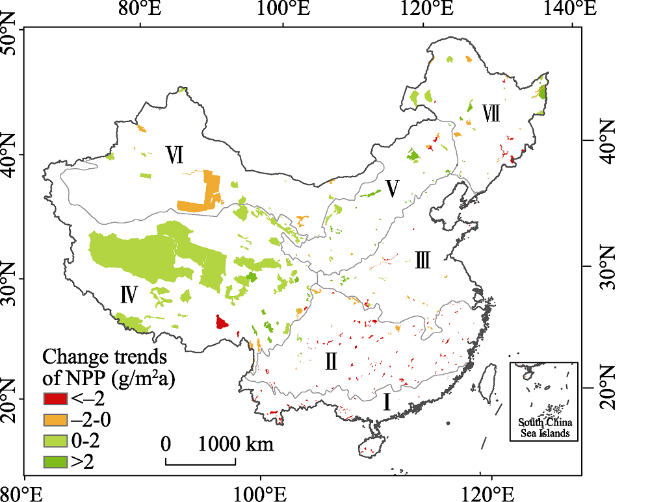
Figure 3 Spatial variations of FVC in typical NNRs from 2000 to 2015 |
Figure 4 The average NPP in typical NNRs (a) with different types and (b) located in varied climate zones |
Figure 5 Spatial variations of NPP in typical NNRs from 2000 to 2015 |
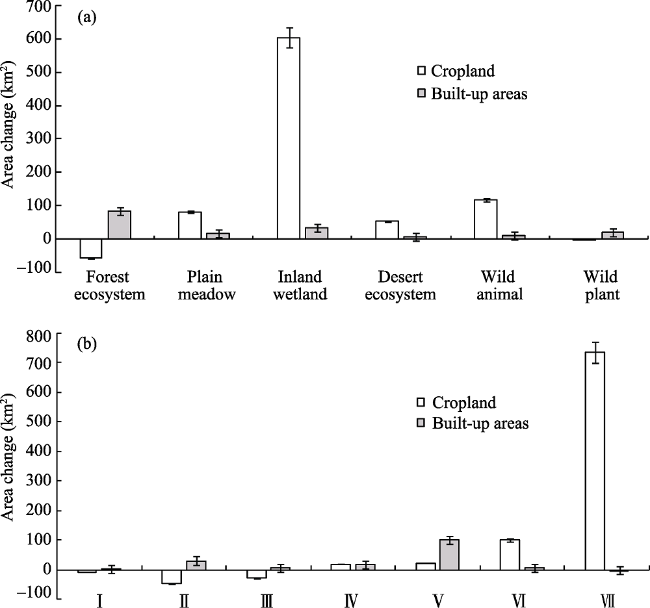
Figure 6 The area changes of cropland and built-up areas in typical NNRs (a) with different types and (b) located in varied climate zones |
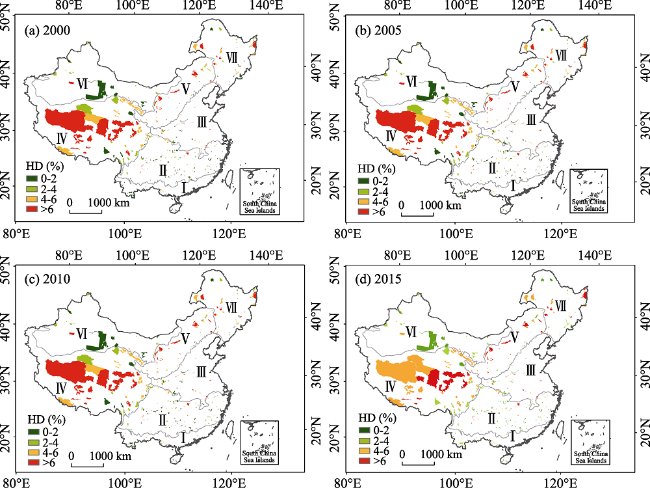
Figure 7 Spatial distribution of HD in typical NNRs for the years of 2000, 2005, 2010 and 2015 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
Millennium Ecosystem Assessment Board, 2003. Ecosystems and human well-being. Washington,DC: Island Press.
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}