
Journal of Geographical Sciences >
The spatiotemporal responses of Populus euphratica to global warming in Chinese oases between 1960 and 2015
Author: Zhang Wenxia (1987-), MS Candidate, specialized in arid area environment and the construction of oasis. E-mail: zwxjiayou@sina.com
*Corresponding author: Liu Puxing (1964-), PhD and Professor, E-mail: liupx2016@163.com
Received date: 2017-09-17
Accepted date: 2017-10-30
Online published: 2018-03-30
Supported by
National Natural Science Foundation of China, No.40961035, No.41461012
The Science and Technology Project of Gansu Province, No.0803RJZA094
Copyright
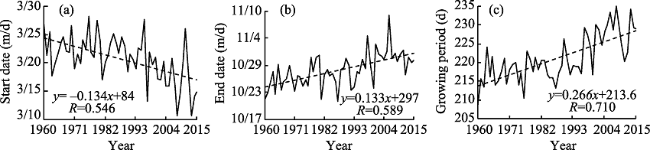
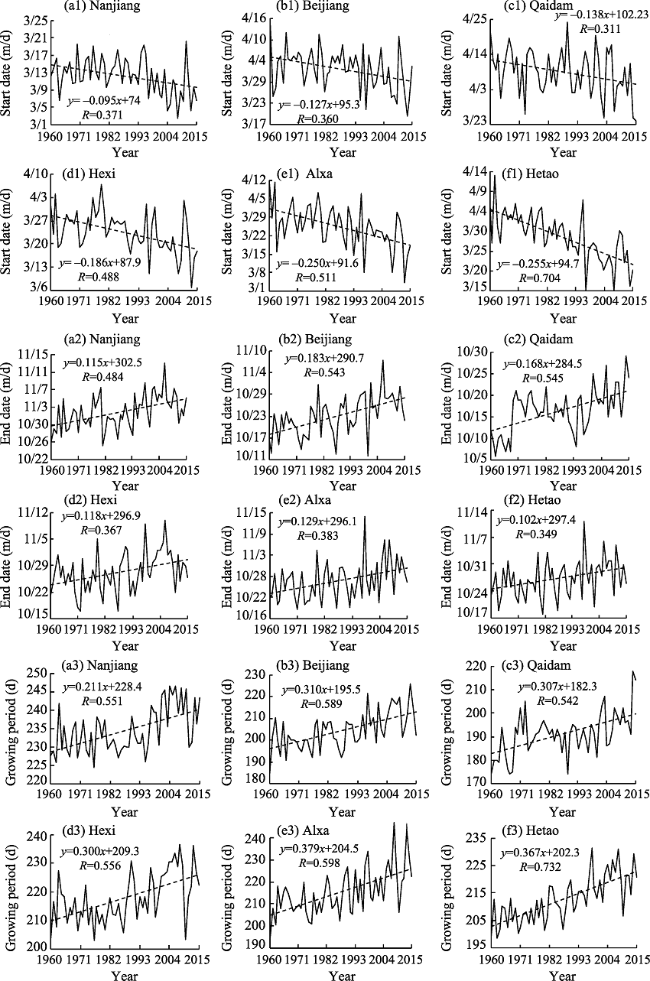
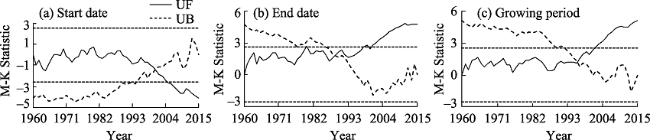
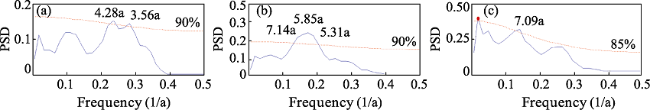
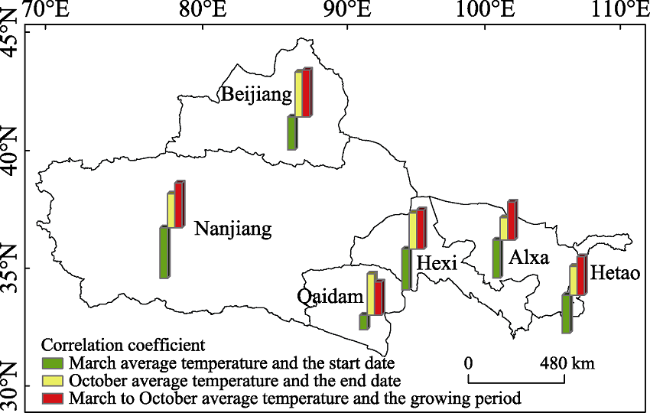
Daily average temperature data from 48 meteorological stations in Chinese oases that are within the distribution area of Populus euphratica were analyzed to determine the spatiotemporal responses of this tree to climate change. Specifically, the start and end date as well as the number of days that comprised the growing season were analyzed with a multi-year trend line and using the Mann-Kendall mutation test, inverse distance weighted interpolation (IDW) in the software ArcGIS, a Morlet wavelet power spectrum, and correlation analysis. The results of this study show that, over the last 56 years, the start date of the P. euphratica growing season has advanced, while the end date has been postponed, and the number of days that comprise the growing season have gradually increased. The changing trend rates recovered in this analysis for these three time slices are -1.34 d/10 a, 1.33 d/10a, and 2.66 d/10a (α ≥ 0.001), respectively. Data show that while spatial disparity is extremely significant, it is nevertheless the case that along a southwest-to-northeast transect of Chinese oases, the later the start date of the P. euphratica season, the sooner the end data and the shorter the growing season. Mutations points in start and end date, as well as for the growing season overall were observed in 2001, 1989, and 1996, respectively, and the data presented in this paper show that, in particular, the date of this end of this period is most sensitive to climate warming. Growing season cycles for P. euphratica are between 3.56 years and 7.14 years, consistent with the periodicity of El Niño events, while a start date cycle between 3.56 years and 4.28 years is consistent with atmospheric circulation cyclicity. The causal analysis presented in this paper shows that the Asian polar vortex area index (APVAI), the Qinghai-Tibet Plateau index (TPI), the westerly circulation index (WCI), and carbon dioxide emissions (CDE) are the main factors influencing spatiotemporal changes in the growth of P. euphratica, the effect of latitude during the growing season is more significant than altitude, and the start date of the growing season is more significantly influenced by these factors than end date. In addition, data show that the start date, end date, and length of the growing season are all significantly correlated with their average corresponding monthly temperature (correlation coefficients are -0.875, 0.770, and 0.897; α≥0.001). Thus, if the average temperature in March increases by 1℃, the start date of the growing season will advance by 2.21 days, while if the average temperature in October increases by the same margin then the seasonal end date will be delayed by 2.76 days. Similarly, if the average temperature between March and October increases by 1℃, the growing season will be extended by 7.78 days. The results of this study corroborate the fact that changes in the P. euphratica growing are sensitive to regional warming and are thus of considerable theoretical significance to our understanding of the responses of Chinese vegetation to climate change as well as to ecological restoration.
ZHANG Wenxia , FENG Qingrong , WANG Tianguang , WANG Tianqiang . The spatiotemporal responses of Populus euphratica to global warming in Chinese oases between 1960 and 2015[J]. Journal of Geographical Sciences, 2018 , 28(5) : 579 -594 . DOI: 10.1007/s11442-018-1492-y
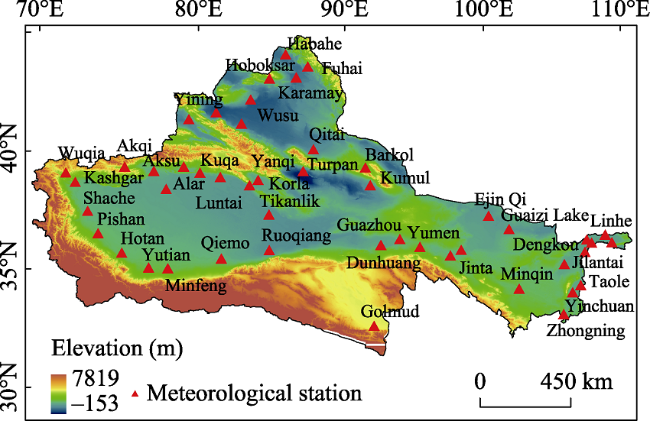
Figure 1 Distribution of meteorological stations in Chinese oases characterized by P. euphratica |
Figure 2 Trends in P. euphratica start date, end date, and growing season variation in Chinese oases |
Figure 3 Trends in P. euphratica start date, end date, and growing season variation in each of the Chinese oases sampled in this study (Abbreviations: Beijiang, northern Xinjiang oasis; Nanjiang, southern Xinjiang oasis; Hexi, Hexi Corridor oasis; Hetao, Hetao Plain oasis; Qaidam, Qaidam Basin oasis; Alxa, Alxa oasis) |
Table 1 Decadal P. euphratica growing season anomalies in Chinese oases |
| Decade | Start date (d) | End date (d) | Growing season (d) |
|---|---|---|---|
| 1960-1969 | 1.8 | -2.7 | -4.4 |
| 1970-1979 | 2.4 | -1.3 | -3.7 |
| 1980-1989 | 1.6 | -1.5 | -3.2 |
| 1990-1999 | -0.1 | 0.5 | 0.7 |
| 2000-2009 | -3.8 | 3.6 | 7.3 |
| 2010-2015 | -3.3 | 2.3 | 5.5 |
Figure 4 Trends in the spatial distribution of the P. euphratica growing season in the Chinese oases sampled in this study |
Figure 5 Mutation analysis of the start date, end date, and growing season of P. euphratica in Chinese oases |
Table 2 Changes in the P. euphratica growing season in Chinese oases before, and after mutation |
| Start date | End date | Growing season (d) | |
|---|---|---|---|
| Before abrupt change | 3/22 | 10/26 | 218 |
| After abrupt change | 3/17 | 10/30 | 228 |
| Change (d) | -5 | 4 | 10 |
Figure 6 Morlet wavelet power spectra of the start date, end date, and the number of days that comprise the P. euphratica growing season in Chinese oases |
Table 3 Correlation analysis results comparing the P. euphratica growing season and relevant impact factors in Chinese oases |
| APVII | APVAI | CA | TPI | WCI | CDE | ||
|---|---|---|---|---|---|---|---|
| Start date | Alxa | 0.243* | 0.529*** | 0.029 | -0.185 | -0.399*** | -0.377*** |
| Beijiang | 0.393*** | 0.583*** | -0.053 | -0.177 | -0.263** | -0.246* | |
| Qaidam | -0.091 | 0.013 | -0.108 | -0.346*** | -0.092 | -0.17 | |
| Hetao | 0.173 | 0.486*** | 0.045 | -0.306** | -0.397*** | -0.582*** | |
| Hexi | 0.181 | 0.580*** | -0.016 | -0.434*** | -0.409*** | -0.319** | |
| Nanjiang | -0.21 | 0.342*** | -0.272** | -0.540*** | -0.435*** | -0.321** | |
| End date | Alxa | -0.104 | -0.273** | -0.163 | 0.372*** | 0.315** | 0.281** |
| Beijiang | -0.324** | -0.523*** | -0.179 | 0.372*** | 0.284** | 0.547*** | |
| Qaidam | -0.340** | -0.042 | 0.071 | 0.135 | 0.068 | 0.474*** | |
| Hetao | 0.145 | -0.211 | -0.096 | 0.395*** | 0.382*** | 0.392*** | |
| Hexi | -0.03 | -0.395*** | -0.122 | 0.581*** | 0.308** | 0.406*** | |
| Nanjiang | -0.230* | -0.416*** | -0.012 | 0.481*** | 0.236* | 0.529*** | |
| Growing season | Alxa | -0.152 | -0.358*** | -0.246* | 0.301** | 0.356*** | 0.507*** |
| Beijiang | -0.205 | -0.368*** | -0.152 | 0.345*** | 0.234* | 0.538*** | |
| Qaidam | -0.393*** | -0.270** | -0.062 | 0.192 | 0.059 | 0.423*** | |
| Hetao | -0.106 | -0.363*** | -0.22 | 0.361*** | 0.414*** | 0.649*** | |
| Hexi | -0.169 | -0.362*** | -0.197 | 0.445*** | 0.335** | 0.488*** | |
| Nanjiang | -0.222* | -0.480*** | 0.041 | 0.592*** | 0.322** | 0.552*** |
Abbreviations: *, ** and, *** denote α = 0.1, α = 0.05, and α = 0.01 levels of significance, respectively. |
Figure 7 Correlation analyses comparing start dates, end dates, and the number of days that comprise the P. euphratica growing season versus mean monthly temperatures in Chinese oases |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}