As a result of continued anthropogenic disturbances, climate change has and will continue to affect the forest ecosystem from species to communities, including tree species richness, forest productivity, forest composition and distribution, and forest biomass (
Boisvenue and Running, 2006;
Bertrand et al., 2011;
Michaletz et al., 2014;
Dai et al., 2016). Among them, the change of forest biomass is the most significant criterion for evaluating the quality of the forests and has been widely used in the studies of forest ecological responses to climate change and other disturbances (
Fearnside, 2000;
Mickler et al., 2002). In general, climate change and its impact on forest ecosystems show a long-term process (
Payette et al., 1989). Much attention has been focused on the interaction between atmospheric composition changes (e.g., COB
2B, N) and the tree species or forest communities (
Medlyn et al., 2001;
Xu et al., 2007). However, compared with the long-term effects caused by climate change, land use change (such as urbanization, deforestation, farmland expansion and transition, and engineering construction) can bring more direct, short-term effects on forest landscape and forest biomass (
Rudel et al., 2005). Due to the complex relationships between forest ecosystem, climate system and land system, it is difficult to explore and predict the key ecosystem characteristics except by going through coarse estimates (
Gustafson et al., 2010). In recent years, however, multi-scale and multi-model approaches have provided a new method for some organizations, communities and researchers to deal with these comprehensive issues. For example, global land project (GLP) presented a framework that incorporates human behavior processes in land use and climate system models to quantitatively analyze dynamic changes in terrestrial ecosystems by using agent-based land use and land cover change models (ABM/LUCC) and some dynamic global vegetation models (DGVMs) (
Rounsevell et al., 2012). And for another example, the European advanced terrestrial ecosystem analysis and modeling (ATEAM) project proposed using multiple internally consistent scenarios and models to assess the vulnerability of agriculture, forestry, and other human sectors that rely on ecosystem services with respect to global change (
Schröter et al., 2004). In terms of more specific issues for forest ecosystems, many studies have also been conducted at the regional or landscape levels. Nepstad
et al. (2008) used global circulation models (GCMs), dynamic vegetation models and economic models to explore the interactions among ecosystems, economy and climate to make prospects for a near-term forest tipping point in the Amazon. Thompson
et al. (2011) used the LANDIS-II model to evaluate regional forest growth and composition change in Massachusetts, USA over a period of 50 years. They found that, while climate change may enhance growth rates, it will be more than offset by land use, primarily by forest conversion for developed use. ABM has many advantages in simulating land use change and it is designed to integrate human decision processes into a location-specific context in order to explain patterns of land use and test the understanding of land use functions (
Matthews et al., 2007). For forest landscape simulation, the LANDIS-II model is a forest disturbance and succession model that can simulate significant ecological characteristics of forested landscapes, such as tree composition, distribution, disturbances, and seed dispersal, as well as the spatial arrangement of aboveground biomass (AGB) (
Scheller et al., 2007). Meanwhile, this model was specifically designed to address the effect of land use and climate change on forest landscape (
Xu et al., 2007,
2011;
Thompson et al., 2016). Based on the above, we combined an ABM/LUCC model, an ecosystem process model (PnET-II) and the LANDIS-II to simulate and analyze the variations in forest AGB under various land use and climate scenarios.

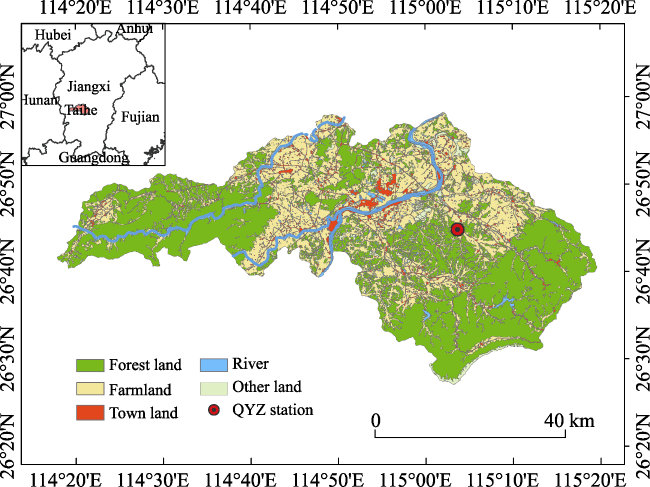
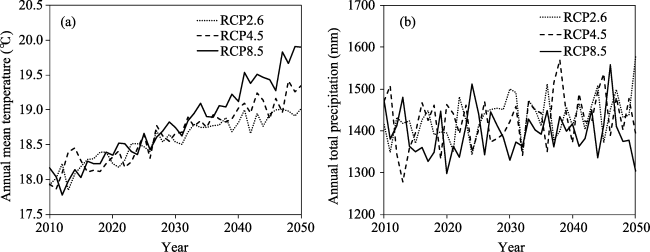
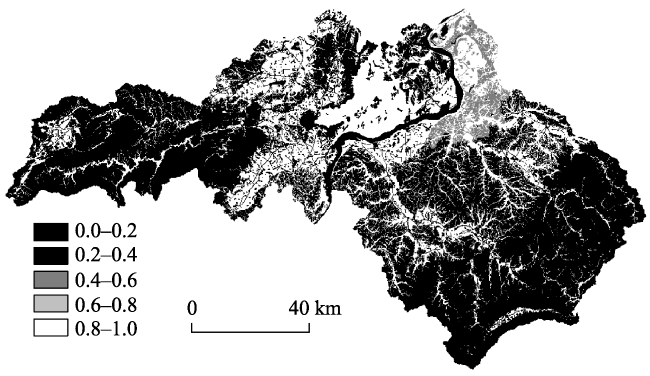
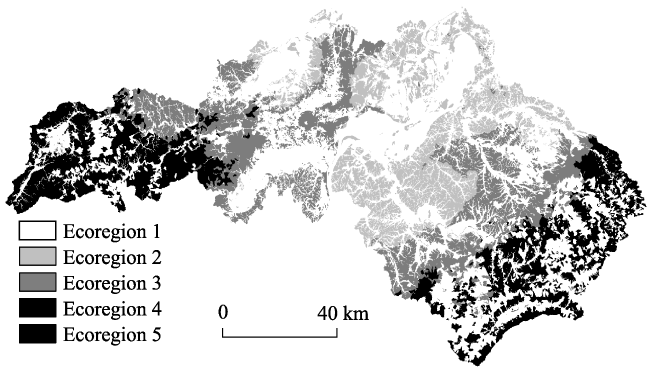
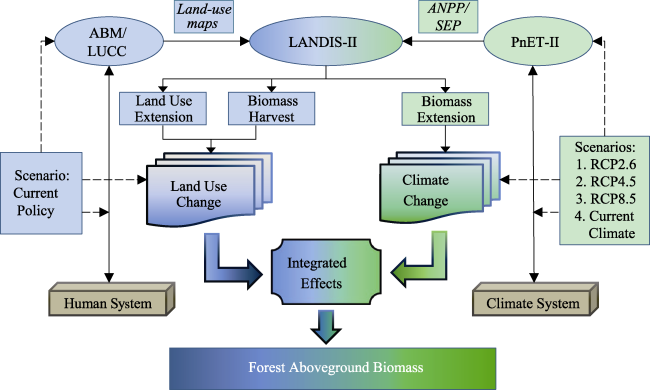
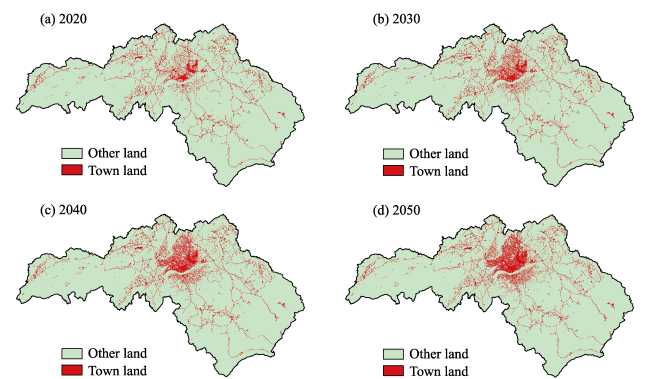
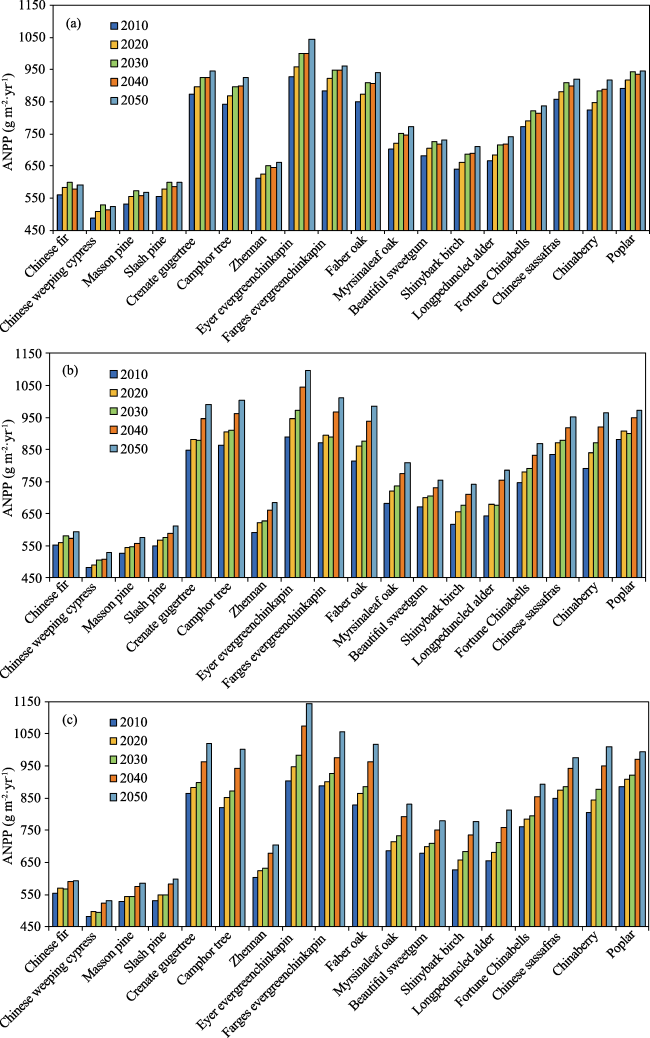
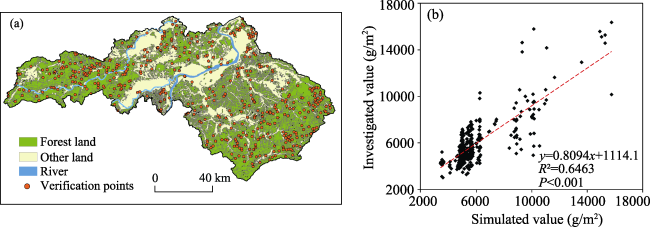
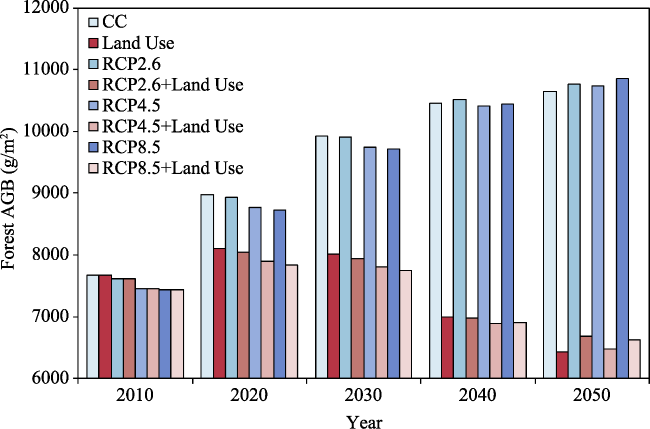
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}