
Journal of Geographical Sciences >
A systematic review of research studies on the estimation of net primary productivity in the Three-River Headwater Region, China
Author: Sun Qingling (1991-), PhD Candidate, specialized in ecological modelling. E-mail: sunql@lreis.ac.cn
*Corresponding author: Li Baolin (1970-), PhD and Professor, specialized in environmental remote sensing and regional ecological modelling. E-mail: libl@lreis.ac.cn
Received date: 2016-06-24
Accepted date: 2016-07-26
Online published: 2017-04-10
Supported by
National Key Research and Development Program of China, No.2016YFC0500205
National Basic Research Program of China (973 Program), No.2015CB954103, No.2015CB954101
Copyright
The Three-River Headwater Region (TRHR), known as the “Water Tower of China”, is an important ecological shelter for national security interests and regional sustainable development activities for many downstream regions in China and a number of Southeast Asian countries. The TRHR is a high-elevation, cold environment with a unique, but typical alpine vegetation system. Net primary productivity (NPP) is a key vegetation parameter and ecological indicator that can reflect both natural environmental changes and carbon budget levels. Given the unique geographical environment and strategic location of the TRHR, many scholars have estimated NPP of the TRHR by using different methods; however, these estimates vary greatly for a number of reasons. To date, there is no paper that has reviewed and assessed NPP estimation studies conducted in the TRHR. Therefore, in this paper, we (1) summarized the related methods and results of NPP estimation in the TRHR in a systematic review of previous research; (2) discussed the suitability of existing methods for estimating NPP in the TRHR and highlighted the most significant challenges; and (3) assessed the estimated NPP results. Finally, developmental directions of NPP estimation in the TRHR were prospected.
SUN Qingling , LI Baolin , ZHOU Chenghu , LI Fei , ZHANG Zhijun , DING Lingling , ZHANG Tao , XU Lili . A systematic review of research studies on the estimation of net primary productivity in the Three-River Headwater Region, China[J]. Journal of Geographical Sciences, 2017 , 27(2) : 161 -182 . DOI: 10.1007/s11442-017-1370-z
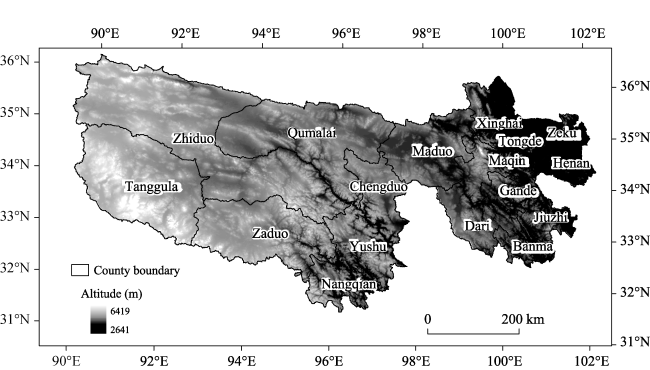
Figure 1 Geographical location and scope of the TRHR |
Table 1 Estimated mean NPP of the TRHR in different studies |
| Study area | Vegetation type | Method & Model | Study period | Mean NPP (g C·m-2 ·yr-1) | Reference |
|---|---|---|---|---|---|
| TRHR | Grassland | Climate model (Thornthwaite Memorial) | 2002-2010 | 481.44 | Li and Zhang, 2014 |
| TRHR | Grassland | Climate model (Production potential model) | 1971-2003 | 225.00 | Li, 2010 |
| TRHR | Grassland | Climate model (Miami) | 2005-2006 | 211.92 | Wang, 2013 |
| TRHR | Grassland | Climate model (CIM) | 2005-2006 | 214.75 | Wang, 2013 |
| TRHR | Grassland | Remote sensing model (CASA) | 2005-2006 | 61.42 | Wang, 2013 |
| TRHR | Grassland | RS-process coupled model (MOD17A3) | 2005-2006 | 93.98 | Wang, 2013 |
| TRHR | Grassland | RS-process coupled model (MOD17A3) | 2000-2010 | 86.80 | Zhang et al., 2015a |
| TRHR | All the vegetation | Climate model (Thornthwaite Memorial) | 1960-2011 | 570.35 | Guo et al., 2013 |
| TRHR | All the vegetation | Climate model (Miami) | 2004-2008 | 486.90 | Cai et al., 2013 |
| TRHR | All the vegetation | Climate model (Zhou Guangsheng) | 2004-2008 | 302.45 | Cai et al., 2013 |
| TRHR | All the vegetation | Climate model (CASA) | 2001-2010 | 169.02 | Zhang et al., 2014 |
| TRHR | All the vegetation | Climate model (CASA) | 2004-2008 | 168.68 | Cai et al., 2013 |
| TRHR | All the vegetation | Remote sensing model (CASA) | 2010 | 146.66 | Wo et al., 2014 |
| TRHR | All the vegetation | Remote sensing model (CASA) | 2003, 2008, 2013 | 148.82 | Chen, 2015 |
| TRHR | All the vegetation | Remote sensing model (GLOPEM) | 1988-2004 | 194.85 | Shao and Fan, 2012 |
| TRHR | All the vegetation | RS-process coupled model (GLOPEM-CEVSA) | 1988-2004 | 143.17 | Wang et al., 2009 |
Table 2 Estimated mean NPP of different vegetation types in the TRHR |
| Vegetation type | Mean NPP (g C·m-2 ·yr-1) | ||||
|---|---|---|---|---|---|
| Shao and Fan, 2012 | Cai et al., 2013 | Wo et al., 2014 | Wang et al., 2009 | Guo et al., 2006 | |
| Grassland | 218.74 | / | 162.87 | 160.90 | / |
| Alpine steppe | / | 129.41 | / | / | 79.34 |
| Alpine meadow | / | 188.95 | / | / | 89.38 |
| Forest | 405.20 | / | 279.81 | 267.90 | / |
| Shrub | 156.04 | / | / | / | / |
| Farmland | 289.22 | / | 256.28 | 222.94 | / |
| Desert | 21.48 | / | 62.81 | 36.13 | / |
| Marsh | 127.09 | / | / | 161.36 | / |
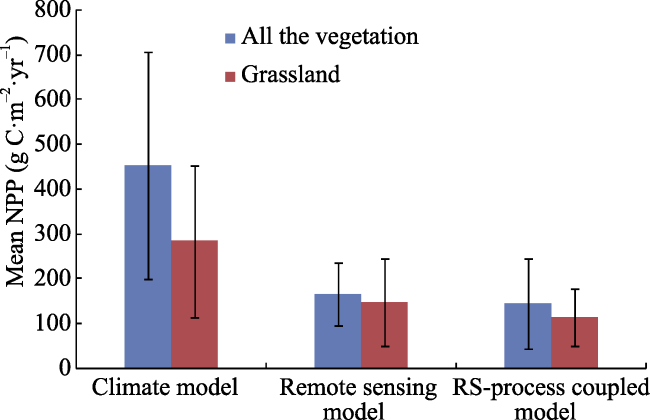
Figure 2 Comparison of estimated NPP based on different models in the TRHR |
Table 3 Measured grassland NPP at different sites in the TRHR |
| Site | Latitude | Longitude | Altitude | Vegetation type | Year | Measured NPP(g C· m-2·yr-1) | Data source |
|---|---|---|---|---|---|---|---|
| Zhenqin | N33°24′30′′ | E97°18′00′′ | 4250 m | Alpine meadow | 2010- 2011 | 118.41 | Fan, 2003 |
| Wudaoliang | N35°12′56′′ | E93°04′05′′ | 4626 m | Alpine steppe | 2000 | 53.55 | Luo et al., 2004 |
| Tuotuohe | N34°18′51′′ | E92°32′52′′ | 4582 m | Alpine steppe | 2000 | 69.30 | Luo et al., 2004 |
| Dawu | N34°23′24′′ | E100°16′33′′ | 3980 m | Alpine meadow | 2014 | 139.07 | Field measurement |
| Maduo | N34°54′40′′ | E98°11′13′′ | 4207 m | Alpine steppe | 2015 | 113.23 | Field measurement |
The original units of NPP in Fan (2003) and Luo et al. (2004) were g DM·m-2·yr-1 and t DM·ha-1·yr-1, respectively. We used 0.45 as the C content to convert dry matter (DM) to C to make these results comparable. |
Table 4 Root to shoot (R/S) ratios of primary grassland types in the TRHR |
| Alpine meadow | Alpine steppe | Temperate steppe | Marsh | Reference | Acquisition mode |
|---|---|---|---|---|---|
| 4.15 | / | / | / | Fan, 2003 | Field measurement |
| 6.8 | 5.2 | / | / | Yang et al., 2009 | Field measurement |
| 7.92 | 4.42 | 4.32 | / | Luo et al., 2002 | Field measurement |
| 7.92 | 4.25 | 4.25 | 15.68 | Piao et al., 2004 | Literature review |
| 9.19 | 9.49 | 9.19 | / | Wang et al., 2008 | Literature review |
| 6.5 | 6.2 | 6.4 | / | Ma et al., 2014 | Literature review |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}