
Journal of Geographical Sciences >
Spatio-temporal analysis of flowering using
LiDAR topography
Received date: 2015-07-15
Accepted date: 2015-10-29
Online published: 2017-02-10
Copyright
Spatio-temporal patterns of flowering in forest ecosystems are hard to quantify and monitor. The objectives of this study were to investigate spatio-temporal patterns (e.g. soils, simple slope classes, slope aspect, and flow accumulation) of flowering around Lake Issaqueena, South Carolina (SC, USA) using plant-flowering database collected with GPS- enabled camera (stored in Picasa 3 web albums and project website) on a monthly basis in 2012 and LiDAR-based topography. Pacolet fine sandy loam had the most flowering plants, followed by Madison sandy loam, both dominant soil types around the lake. Most flowering plants were on moderately steep (17%-30%) and gently sloping (4%-8%) slopes. Most flowering plants were on west (247.5°-292.5°), southwest (202.5°-247.5°), and northwest (292.5°-337.5°) aspects. Most flowering plants were associated with minimum and maximum flows within the landscape. Chi-square tests indicated differences in the distributions of the proportions of flowering plants were significant by soil type, slope, aspect, and flow accumulation for each month (February-November), for all months (overall), and across months. The Chi-square test on area-normalized data indicated significant differences for all months and individual differences by each month with some months not statistically significant. Cluster analysis on flowering counts for nine plant families with the most flowering counts indicated no unique separation by cluster, but implied that the majority of these families were flowering on strongly sloping (9%-16%) slopes, on southwest (202.5°-247.5°) aspects, and low flow accumulation (0-200). Presented methodology can serve as a template for future efforts to quantify spatio-temporal patterns of flowering and other phenological events.
Key words: aspect; flow accumulation; Geographic Information Systems (GIS); phenology; soils
HART Samantha
,
MIKHAILOVA Elena
,
POST Christopher
,
McMILLAN Patrick
,
SHARP Julia
,
BRIDGES William
. Spatio-temporal analysis of flowering using
LiDAR topography[J]. Journal of Geographical Sciences, 2017
, 27(1)
: 62
-78
.
DOI: 10.1007/s11442-017-1364-x
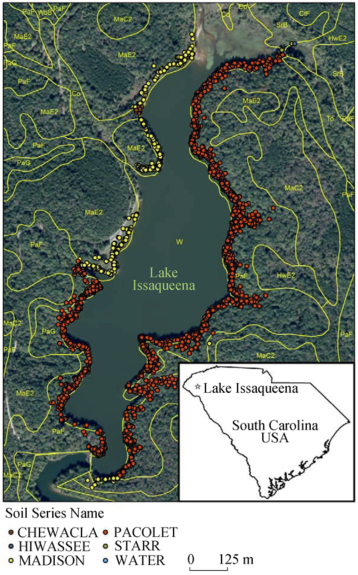
Figure 1 Soil types and flowering occurrences around Lake Issaqueena, SC |
Table 1 Monthly total precipitation (cm) and monthly average temperature (°C) for 2012, and 50-year mean (Source: U.S. Historical Climatology Network-Monthly Data, Site 381770, Clemson University, South Carolina) |
| 2012 | 50-year mean | |||||
|---|---|---|---|---|---|---|
| Month | Mean temp., °C | Precip., cm | Mean temp.,°C | Precip., cm | ||
| January | 9 | 11 | 5 | 13 | ||
| February | 9 | 5 | 7 | 12 | ||
| March | 17 | 6 | 11 | 14 | ||
| April | 18 | 6 | 16 | 10 | ||
| May | 22 | 8 | 20 | 10 | ||
| June | 24 | 16 | 24 | 10 | ||
| July | 27 | 12 | 26 | 11 | ||
| August | 25 | 21 | 25 | 12 | ||
| September | 22 | 6 | 22 | 10 | ||
| October | 16 | 7 | 16 | 10 | ||
| November | 10 | 2 | 11 | 10 | ||
| December | 9 | 13 | 7 | 12 | ||
| Total precip. | 112 | 134 | ||||
| Mean temp. | 17 | 16 | ||||
Table 2 Data sources and descriptions |
| Data layer | Source | Coordinate system | Spatial resolution | Date |
|---|---|---|---|---|
| DEM (LiDAR) | Pickens County GIS | NAD State Plane 1983 SC | 3.048 m | 2011 |
| Lake Polygon | NHD USGS | NAD State Plane 1983 SC | na | 2013 |
| NAIP Aerial Photo | USDA-NRCS | NAD State Plane 1983 SC | 1 m | 2013 |
| SSURGO Soils Data | USDA-NRCS | Geographic | na | na |
Table 3 Flowering counts and area (m2 and %) by soil type around Lake Issaqueena, SC in 2012 |
 |
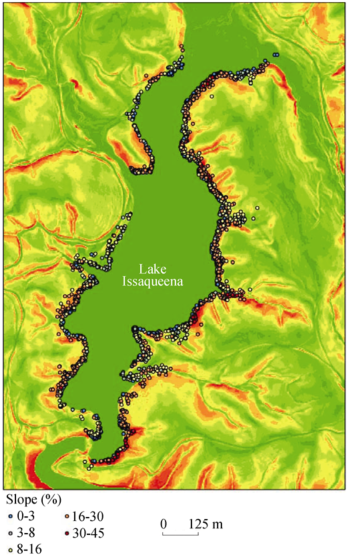
Figure 2 Simple slope classes and flowering occurrences around Lake Issaqueena, SC |
Table 4 Flowering counts area(m2 and %) by simple slope classes (Soil Survey Manual,2015) around Lake Issaqueena, Sc in 2012 |
 |
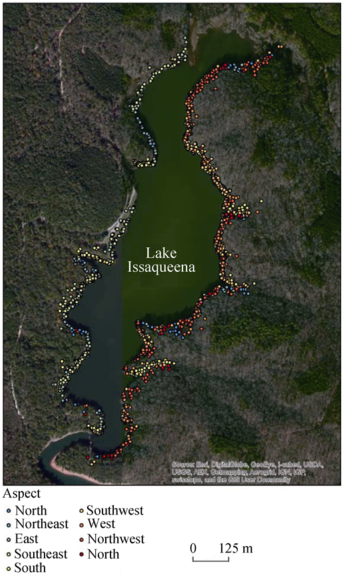
Figure 3 Slope aspect and flowering occurrences around Lake Issaqueena, SC |
Table 5 Flowering counts and area(m2 and %) by slope aspect around Lake Issaqueena, Sc in 2012 |
 |
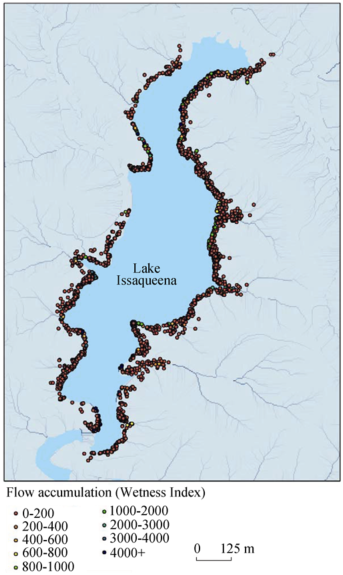
Figure 4 Flow accumulation and flowering occurrences around Lake Issaqueena, SC |
Table 6 Flowering counts and area (m2 and %) by flow accumulation around Lake Issaqueena, Sc in 2012 |
 |
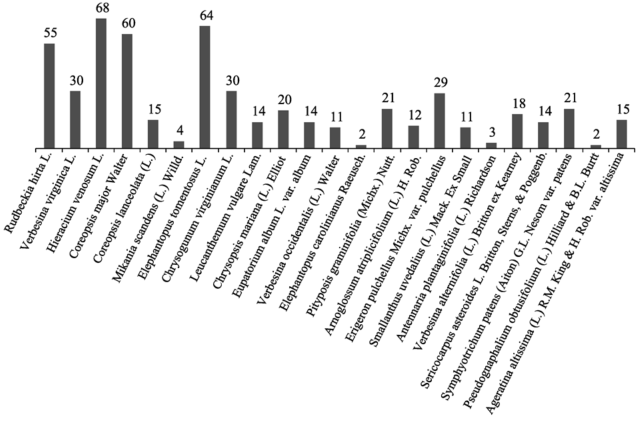
Figure 5 Flowering counts distribution within the Asteraceae family |
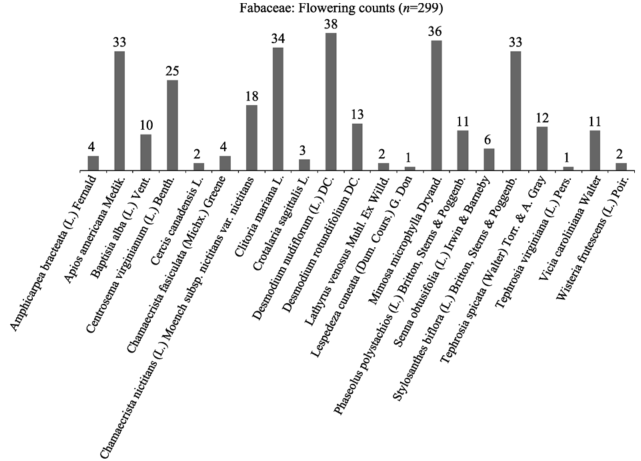
Figure 6 Flowering counts distribution within the Fabaceae family |
Table 7 Landscape characteristics associated with the top nine families in terms of flowering counts around Lake Issaqueena, SC in 2012 |
| Family | Slope aspect | Slope | Flow accumulation | ||||
|---|---|---|---|---|---|---|---|
| n | Mean | St dev. | Mean | St dev. | Mean | St dev. | |
| Overall | |||||||
| Asteraceae | 533 | 215.3 | 80.7 | 16 | 8 | 453 | 3392 |
| Campanulaceae | 97 | 180.0 | 97.8 | 14 | 10 | 223 | 777 |
| Caryophyllaceae | 82 | 229.7 | 93.5 | 19 | 8 | 448 | 1617 |
| Clusiaceae | 84 | 218.3 | 86.9 | 13 | 8 | 218 | 737 |
| Ericaceae | 105 | 249.1 | 80.6 | 18 | 11 | 293 | 1229 |
| Fabaceae | 304 | 183.6 | 82.7 | 16 | 8 | 758 | 6260 |
| Lamiaceae | 101 | 231.6 | 61.3 | 17 | 8 | 97 | 510 |
| Liliaceae | 132 | 243.4 | 94.9 | 17 | 8 | 79 | 276 |
| Melastomataceae | 115 | 192.7 | 78.5 | 11 | 8 | 2816 | 7502 |
| February | |||||||
| Asteraceae | 2 | 167.0 | 0.0 | 17 | 0 | 1 | 0 |
| March | |||||||
| Asteraceae | 52 | 212.9 | 88.9 | 18 | 9 | 103 | 257 |
| Caryophyllaceae | 26 | 210.6 | 83.7 | 18 | 8 | 104 | 313 |
| Ericaceae | 21 | 192.9 | 119.0 | 11 | 6 | 333 | 1003 |
| Fabaceae | 22 | 183.4 | 110.3 | 16 | 9 | 229 | 725 |
| Lamiaceae | 4 | 222.4 | 2.2 | 15 | 3 | 9 | 0 |
| Liliaceae | 36 | 222.8 | 116.0 | 16 | 6 | 46 | 185 |
| April | |||||||
| Asteraceae | 61 | 270.8 | 42.0 | 21 | 8 | 287 | 1395 |
| Caryophyllaceae | 10 | 265.9 | 33.2 | 17 | 7 | 1060 | 2308 |
| Ericaceae | 66 | 274.4 | 52.1 | 20 | 10 | 341 | 1443 |
| Fabaceae | 7 | 275.3 | 60.3 | 18 | 10 | 142 | 247 |
| Lamiaceae | 12 | 257.6 | 87.7 | 17 | 9 | 417 | 1393 |
| Liliaceae | 52 | 238.1 | 108.1 | 13 | 7 | 50 | 161 |
| May | |||||||
| Asteraceae | 64 | 224.4 | 80.8 | 13 | 8 | 231 | 695 |
| Campanulaceae | 8 | 125.2 | 21.3 | 27 | 4 | 9 | 4 |
| Caryophyllaceae | 12 | 309.3 | 34.6 | 22 | 10 | 950 | 3155 |
| Ericaceae | 7 | 257.1 | 19.4 | 23 | 7 | 5 | 7 |
| Fabaceae | 52 | 175.8 | 73.7 | 17 | 7 | 198 | 719 |
| Lamiaceae | 32 | 249.1 | 45.5 | 17 | 9 | 81 | 257 |
| Liliaceae | 8 | 283.4 | 38.2 | 18 | 8 | 313 | 378 |
| Melastomataceae | 6 | 123.4 | 36.1 | 7 | 8 | 2253 | 3092 |
| June | |||||||
| Asteraceae | 62 | 222.7 | 78.9 | 17 | 7 | 146 | 516 |
| Campanulaceae | 21 | 227.7 | 101.4 | 15 | 11 | 52 | 102 |
| Caryophyllaceae | 4 | 60.5 | 85.5 | 27 | 2 | 1 | 1 |
| Clusiaceae | 14 | 191.0 | 77.6 | 12 | 11 | 21 | 17 |
| Ericaceae | 3 | 181.7 | 138.4 | 25 | 3 | 38 | 9 |
| Fabaceae | 46 | 218.4 | 76.8 | 16 | 7 | 163 | 699 |
| Lamiaceae | 9 | 231.6 | 71.1 | 17 | 8 | 122 | 336 |
| Liliaceae | 16 | 244.5 | 43.4 | 20 | 10 | 6 | 6 |
| Melastomataceae | 28 | 215.6 | 79.0 | 11 | 7 | 5330 | 12178 |
| July | |||||||
| Asteraceae | 121 | 194.3 | 76.2 | 17 | 7 | 237 | 1541 |
| Campanulaceae | 11 | 163.4 | 135.8 | 9 | 5 | 3 | 2 |
| Caryophyllaceae | 22 | 232.0 | 101.2 | 18 | 7 | 417 | 1321 |
| Clusiaceae | 49 | 221.2 | 88.3 | 14 | 7 | 358 | 944 |
| Ericaceae | 5 | 244.8 | 37.0 | 18 | 15 | 11 | 4 |
| Fabaceae | 78 | 192.9 | 80.9 | 15 | 8 | 199 | 726 |
| Lamiaceae | 27 | 219.6 | 58.2 | 20 | 8 | 18 | 54 |
| Liliaceae | 8 | 264.4 | 49.2 | 30 | 3 | 6 | 5 |
| Melastomataceae | 52 | 187.6 | 81.7 | 11 | 9 | 2094 | 5130 |
| August | |||||||
| Asteraceae | 84 | 203.4 | 79.6 | 13 | 8 | 610 | 2147 |
| Campanulaceae | 43 | 161.2 | 90.0 | 13 | 9 | 419 | 1125 |
| Caryophyllaceae | 7 | 195.5 | 91.0 | 18 | 9 | 405 | 844 |
| Clusiaceae | 21 | 229.6 | 89.9 | 13 | 7 | 21 | 40 |
| Fabaceae | 89 | 162.0 | 78.0 | 14 | 8 | 2145 | 11447 |
| Lamiaceae | 16 | 199.1 | 62.8 | 13 | 6 | 40 | 93 |
| Liliaceae | 12 | 286.0 | 16.2 | 18 | 8 | 295 | 692 |
| Melastomataceae | 29 | 193.8 | 71.4 | 11 | 8 | 1802 | 5295 |
| September | |||||||
| Asteraceae | 36 | 195.1 | 92.5 | 14 | 8 | 1219 | 4083 |
| Campanulaceae | 9 | 221.6 | 75.1 | 14 | 5 | 8 | 7 |
| Caryophyllaceae | 1 | 273.7 | NA | 8 | NA | 39 | NA |
| Fabaceae | 8 | 122.3 | 15.7 | 26 | 4 | 12 | 8 |
| Lamiaceae | 1 | 236.5 | NA | 18 | NA | 1 | NA |
| October | |||||||
| Asteraceae | 37 | 224.3 | 72.0 | 17 | 9 | 1912 | 11277 |
| Campanulaceae | 5 | 190.6 | 91.4 | 22 | 9 | 471 | 420 |
| Ericaceae | 2 | 97.4 | 5.8 | 0 | 0 | 528 | 152 |
| Fabaceae | 2 | 104.9 | 31.6 | 11 | 9 | 2 | 2 |
Table 8 Cluster analysis for landscape characteristics associated with the top nine families in terms of flowering counts around Lake Issaqueena, SC in 2012 |
| Cluster | |||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| Family | |||||
| Asteraceae | 8 | 14 | 510 | 0 | 1 |
| Campanulaceae | 0 | 5 | 92 | 0 | 0 |
| Caryophyllaceae | 1 | 5 | 76 | 0 | 0 |
| Clusiaceae | 0 | 3 | 81 | 0 | 0 |
| Ericaceae | 2 | 3 | 100 | 0 | 0 |
| Fabaceae | 3 | 12 | 282 | 1 | 1 |
| Lamiaceae | 0 | 1 | 100 | 0 | 0 |
| Liliaceae | 0 | 1 | 131 | 0 | 0 |
| Melastomataceae | 12 | 9 | 91 | 3 | 0 |
| Parameter | |||||
| Slope Aspect (°) | 179 | 141 | 215 | 262 | 198 |
| Simple Slope (%) | 5 | 7 | 16 | 2 | 1 |
| Flow Accumulation | 12870 | 3260 | 57 | 41257 | 81820 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
ESRI, 2013. ArcGIS Desktop: Release 10.2 Redlands, CA: Environmental Systems Research Institute.
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
Google, Inc., 2010. Picasa Web Albums Data API for .NET (Version 1.0). Mountain View, CA. Available at
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
SAS v. 9.3, 2011. Copyright, SAS Institute Inc. SAS and all other SAS Institute Inc. product or service names are registered trademarks or trademarks of SAS Institute Inc., , Cary, NCUSA.
|
| [28] |
|
| [29] |
|
| [30] |
USDA, NRCS, 2014. The PLANTS Database (, 28 January 2014). National Plant Data Team, Greensboro, NC 27401-4901 USA.
|
| [31] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}