
Journal of Geographical Sciences >
Latitudinal variation of leaf morphological traits from species to communities along a forest transect in eastern China
Author: Wang Ruili (1988-), PhD, specialized in variation of plant functional traits. E-mail: loverain.88@163.com
*Corresponding author: Yu Guirui, Professor, E-mail: yugr@igsnrr.ac.cn.He Nianpeng, Associate Professor, E-mail: henp@igsnrr.ac.cn
Received date: 2015-08-21
Accepted date: 2015-09-22
Online published: 2016-01-25
Supported by
National Natural Science Foundation of China, No.31290221, No.31470506
Chinese Academy of Sciences Strategic Priority Research Program, No.XDA05050702
Program for Kezhen Distinguished Talents in Institute of Geographic Sciences and Natural Resources Research, CAS, No.2013RC102
Copyright
Comprehensive information on geographic patterns of leaf morphological traits in Chinese forests is still scarce. To explore the spatial patterns of leaf traits, we investigated leaf area (LA), leaf thickness (LT), specific leaf area (SLA), and leaf dry matter content (LDMC) across 847 species from nine typical forests along the North-South Transect of Eastern China (NSTEC) between July and August 2013, and also calculated the community weighted means (CWM) of leaf traits by determining the relative dominance of each species. Our results showed that, for all species, the means (± SE) of LA, LT, SLA, and LDMC were 2860.01 ± 135.37 mm2, 0.17 ± 0.003 mm, 20.15 ± 0.43 m2 kg-1, and 316.73 ± 3.81 mg g-1, respectively. Furthermore, latitudinal variation in leaf traits differed at the species and community levels. Generally, at the species level, SLA increased and LDMC decreased as latitude increased, whereas no clear latitudinal trends among LA or LT were found, which could be the result of shifts in plant functional types. When scaling up to the community level, more significant spatial patterns of leaf traits were observed (R2 = 0.46-0.71), driven by climate and soil N content. These results provided synthetic data compilation and analyses to better parameterize complex ecological models in the future, and emphasized the importance of scaling-up when studying the biogeographic patterns of plant traits.
WANG Ruili , *YU Guirui , *HE Nianpeng , WANG Qiufeng , ZHAO Ning , XU Zhiwei . Latitudinal variation of leaf morphological traits from species to communities along a forest transect in eastern China[J]. Journal of Geographical Sciences, 2016 , 26(1) : 15 -26 . DOI: 10.1007/s11442-016-1251-x
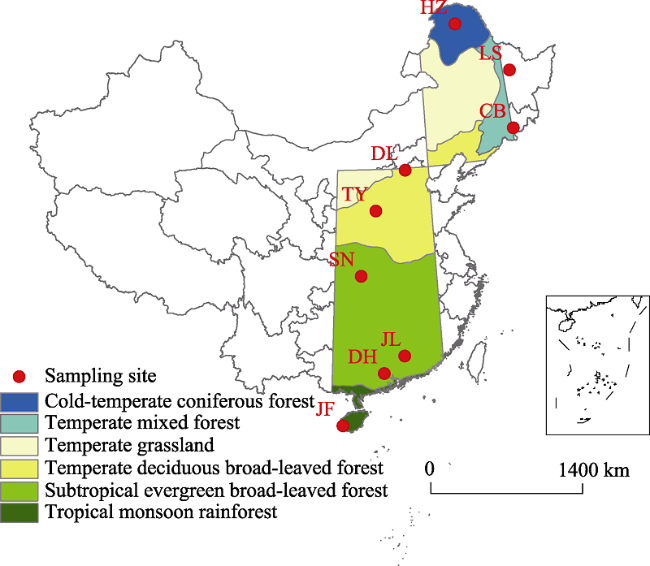
Figure 1 Geographic locations and vegetation types of sampling sites. JF, Jianfengling; DH, Dinghu Mountain; JL, Jiulian Mountain; SN, Shennongjia; TY, Taiyue Mountain; DL, Dongling Mountain; CB, Changbai Mountain; LS, Liangshui; HZ, Huzhong. Different colors highlighted in the North-South Transect of Eastern China (NSTEC) represent different vegetation types (Zhang and Yang, 1995). |
Table 1 Environmental characteristics and vegetation types of sampling sites |
| Site | Latitude (°N) | Longitude (°E) | MAT (°C) | MAP (mm) | SN (mg g-1) | Vegetation type | No. of species |
|---|---|---|---|---|---|---|---|
| JF | 18.7 | 108.9 | 19.8 | 2449.0 | 1.95 | Tropical monsoon rainforest | 139 |
| DH | 23.2 | 112.5 | 20.9 | 1927.0 | 1.76 | Subtropical evergreen broad-leaved forest | 158 |
| JL | 24.6 | 114.4 | 16.7 | 1954.0 | 2.35 | Subtropical evergreen broad-leaved forest | 172 |
| SN | 31.3 | 110.5 | 10.6 | 1330.0 | 3.76 | Subtropical mixed evergreen and deciduous broad-leaved forest | 120 |
| TY | 36.7 | 112.1 | 6.2 | 662.0 | 2.56 | Temperate deciduous broad-leaved forest | 76 |
| DL | 40.0 | 115.4 | 4.8 | 539.1 | 3.12 | Temperate deciduous broad-leaved forest | 79 |
| CB | 42.4 | 128.1 | 2.6 | 691.0 | 6.37 | Temperate mixed forest | 109 |
| LS | 47.2 | 128.9 | -0.3 | 676.0 | 4.59 | Temperate mixed forest | 104 |
| HZ | 51.8 | 123.0 | -4.4 | 481.6 | 3.15 | Cold-temperate coniferous forest | 88 |
Note: MAT, mean annual temperature; MAP, mean annual precipitation; SN, soil N content. |
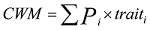
Table 2 Statistics for leaf traits at the species and community levels |
| Level | Traits | n | Mean | Minimum | Maximum | SE | CV | Skewness |
|---|---|---|---|---|---|---|---|---|
| Species | LA (mm2) | 847 | 2860.01 | 4.09 | 56085.4 | 135.37 | 1.51 | 5.54 |
| LT (mm) | 847 | 0.17 | 0.01 | 0.78 | 0.003 | 0.56 | 2.71 | |
| SLA (m2·kg-1) | 847 | 20.15 | 1.89 | 94.99 | 0.43 | 0.68 | 1.29 | |
| LDMC (mg·g-1) | 847 | 316.73 | 44.46 | 775.68 | 3.81 | 0.39 | 0.27 | |
| Community | LACWM (mm2) | 32 | 1443.8 | 22.98 | 3547.5 | 169.35 | 0.66 | 0.49 |
| LTCWM (mm) | 32 | 0.34 | 0.18 | 0.69 | 0.03 | 0.46 | 1.00 | |
| SLACWM (m2·kg-1) | 32 | 9.83 | 5.08 | 18.34 | 0.71 | 0.41 | 0.68 | |
| LDMCCWM (mg·g-1) | 32 | 421.78 | 364.16 | 544.01 | 8.76 | 0.12 | 0.72 |
Note: n, number of species or plots; SE, standard error; CV, coefficient of variation. |
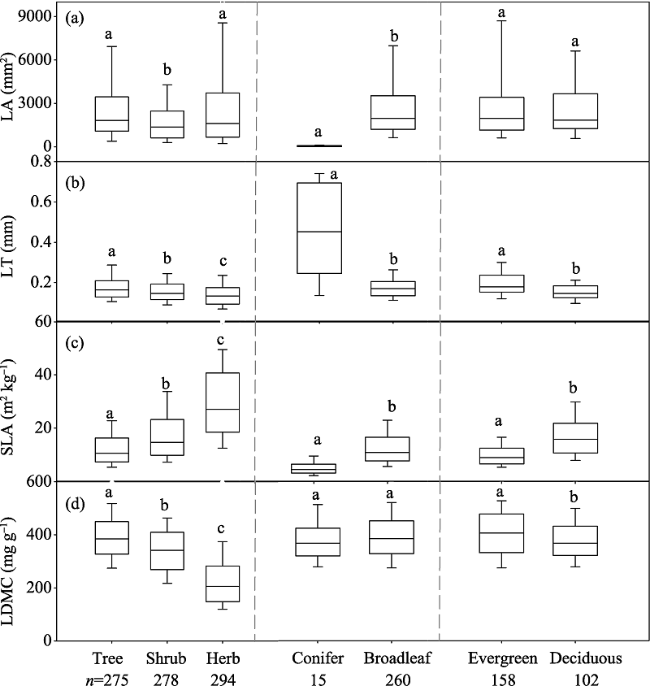
Figure 2 Differences in leaf traits among plant functional types. The black lines across the boxes are median values. n, species number. Statistical differences are denoted using different letters (P < 0.05). |
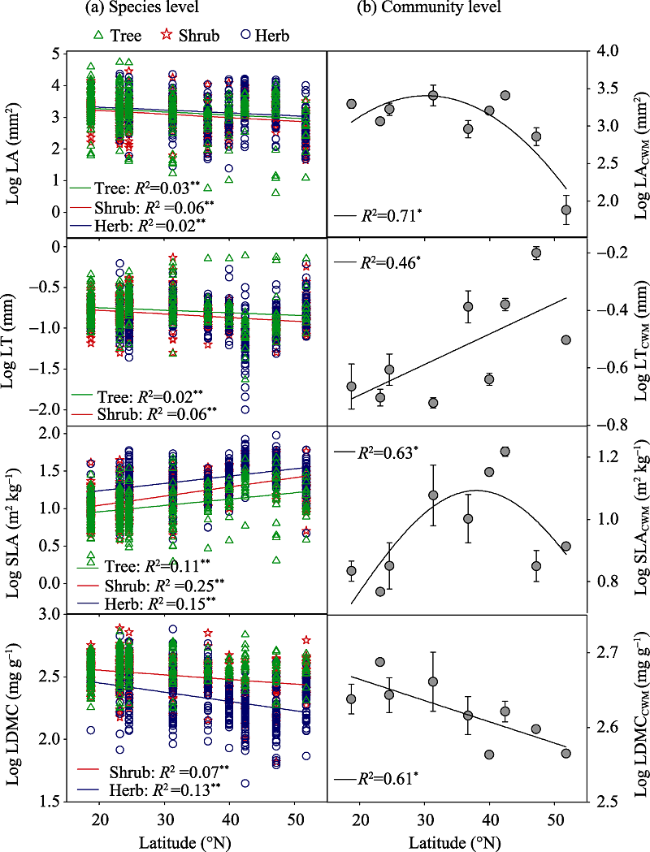
Figure 3 Latitudinal patterns of leaf traits at the species (a) and community levels (b). Error bars in panels (b) represent ± 1 standard error. Only significant regressions are given (P < 0.05). |
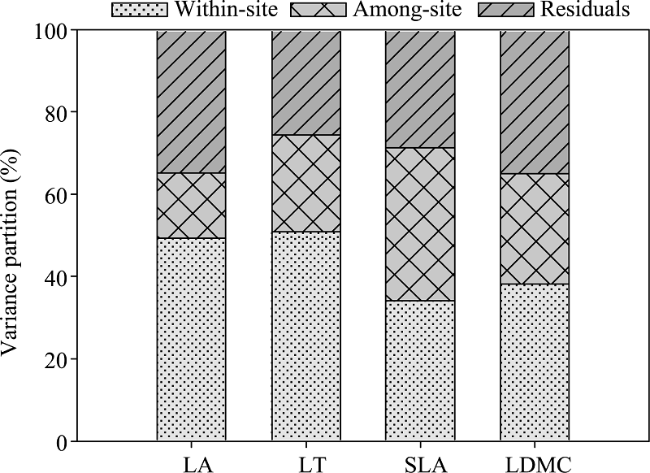
Figure 4 Variance partitioning of species-level leaf traits into within-site, among-site, and residual components |
Table 3 Influences of the plant functional type and environmental factors on species-level leaf traits |
| Factor | Log LA | Log LT | Log SLA | Log LDMC | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | SS% | df | F | SS% | df | F | SS% | df | F | SS% | |
| PFT | 4 | 56.79** | 19.43 | 4 | 1.26** | 41.57 | 4 | 163.24** | 36.83 | 4 | 129.50** | 34.18 |
| MAT | 1 | 98.16** | 6.72 | 1 | 41.41** | 2.19 | ||||||
| MAP | 1 | 25.22** | 1.65 | 1 | 127.55** | 5.76 | ||||||
| SN | 1 | 30.49** | 2.09 | 1 | 139.73** | 9.13 | 1 | 197.69** | 8.92 | 1 | 77.47** | 4.09 |
| PFT×MAT | 4 | 9.90** | 2.09 | |||||||||
| PFT×MAP | 4 | 4.45** | 1.16 | 4 | 6.73** | 1.21 | ||||||
| PFT×SN | 4 | 3.90** | 1.02 | 4 | 2.67* | 0.48 | 4 | 4.30** | 0.91 | |||
| MAT×SN | 1 | 10.18** | 0.54 | |||||||||
| MAP×SN | 1 | 80.67** | 5.27 | 1 | 12.48** | 0.56 | ||||||
| Site | 8 | 6.31** | 3.46 | 8 | 5.14** | 2.68 | 8 | 3.86** | 1.39 | 8 | 4.51** | 1.90 |
| Residuals | 997 | 68.31 | 1001 | 65.51 | 992 | 44.85 | 1022 | 54.11 | ||||
Note: PFT, plant functional type; MAT, mean annual temperature; MAP, mean annual precipitation; SN, soil N concentration; df, degrees of freedom; SS%, percentage of sum of squares explained. *, P < 0.05; **, P < 0.01. |
Table 4 Influences of environmental factors on community-level leaf traits |
| Factor | Log LACWM | Log LTCWM | Log SLACWM | Log LDMCCWM | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | SS% | df | F | SS% | df | F | SS% | df | F | SS% | |
| MAT | 1 | 3.34* | 25.08 | 1 | 7.77* | 16.51 | 1 | 22.56** | 48.04 | |||
| MAP | 1 | 5.30** | 32.75 | |||||||||
| SN | 1 | 7.75** | 36.41 | |||||||||
| MAT×SN | 1 | 0.003 | 1.79 | |||||||||
| Site | 8 | 14.09** | 58.43 | 8 | 14.99** | 54.87 | 8 | 5.86* | 15.42 | 8 | 1.471 | 3.01 |
| Residuals | 22 | 16.49 | 22 | 12.38 | 20 | 29.87 | 22 | 48.95 | ||||
Note: MAT, mean annual temperature; MAP, mean annual precipitation; SN, soil N concentration; df, degrees of freedom; SS%, percentage of sum of squares explained. *, P < 0.05; **, P < 0.01. |
The authors have declared that no competing interests exist.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}