
Journal of Geographical Sciences >
Analysis of spatio-temporal features of a carbon source/sink and its relationship to climatic factors in the Inner Mongolia grassland ecosystem
Author: Dai Erfu (1972-), PhD and Professor, specialized in comprehensive study of physical geography, climate change and regional response, simulation of LUCC. E-mail:daief@igsnrr.ac.cn
*Corresponding author: Wu Zhuo (1988-), PhD Candidate, specialized in land use and ecological process. E-mail:wuz.14b@igsnrr.ac.cn
Received date: 2015-09-20
Accepted date: 2015-10-23
Online published: 2016-07-25
Supported by
National Basic Research Program of China (973 Program), No.2015CB452702, No.2012CB416906
National Natural Science Foundation of China, No.41571098, No.41371196
National Key Technology R&D Program, No.2013BAC03B04
Copyright
Global climate change has become a major concern worldwide. The spatio-temporal characteristics of net ecosystem productivity (NEP), which represents carbon sequestration capacity and directly describes the qualitative and quantitative characteristics of carbon sources/sinks (C sources/sinks), are crucial for increasing C sinks and reducing C sources. In this study, field sampling data, remote sensing data, and ground meteorological observation data were used to estimate the net primary productivity (NPP) in the Inner Mongolia grassland ecosystem (IMGE) from 2001 to 2012 using a light use efficiency model. The spatio-temporal distribution of the NEP in the IMGE was then determined by estimating the NPP and soil respiration from 2001 to 2012. This research also investigated the response of the NPP and NEP to the main climatic variables at the spatial and temporal scales from 2001 to 2012. The results showed that most of the grassland area in Inner Mongolia has functioned as a C sink since 2001 and that the annual carbon sequestration rate amounts to 0.046 Pg C/a. The total net C sink of the IMGE over the 12-year research period reached 0.557 Pg C. The carbon sink area accounted for 60.28% of the total grassland area and the sequestered 0.692 Pg C, whereas the C source area accounted for 39.72% of the total grassland area and released 0.135 Pg C. The NPP and NEP of the IMGE were more significantly correlated with precipitation than with temperature, showing great potential for C sequestration.
Key words: Inner Mongolia; grassland ecosystem; NPP; NEP; carbon source; carbon sink
DAI Erfu , HUANG Yu , Wu Zhuo , ZHAO Dongsheng . Analysis of spatio-temporal features of a carbon source/sink and its relationship to climatic factors in the Inner Mongolia grassland ecosystem[J]. Journal of Geographical Sciences, 2016 , 26(3) : 297 -312 . DOI: 10.1007/s11442-016-1269-0
Table 1 Eco-geographical zonal systems of Inner Mongolia |
| Temperature zones | Humid and arid zones | Codes | Eco-zones | Vegetation type |
|---|---|---|---|---|
| I Cold tem- perate zone | A (humid zone) | IA1 | Deciduous coniferous forest north of Greater Hinggan Mts. | Coniferous forest, wetland |
| II Temperate zone | A (humid zone) | IIA3 | Mixed forest east of Songliao Plain | Mixed forest, wetland |
| B (semi-humid zone) | IIB1 | Forest steppe at center of Songliao Plain | Meadow steppe, farmland | |
| IIB2 | Steppe forest at the center of Greater Hinggan Mts. | Broadleaf forest, meadow | ||
| IIB3 | Forest steppe west of Greater Hinggan Mts. | Meadow steppe | ||
| C (semi-arid zone) | IIC1 | Steppe zone in Xiliao River Plain | Typical steppe, farmland | |
| IIC2 | Forest steppe south of Greater Hinggan Mts. | Typical steppe, shrub | ||
| IIC3 | Steppe zone east of Inner Mongolia | Typical steppe, farmland | ||
| IIC4 | Steppe zone of Hulun Buir | Typical steppe, meadow | ||
| D (arid zone) | IID1 | Desert steppe zone in Ordos and west of Inner Mongolia | Typical steppe, desert | |
| IID2 | Desert zone in Alax and Hexi Corridor | Desert, desert steppe | ||
| III Warm temperate zone | B (semi-humid zone) | IIIB3 | Montane deciduous broadleaf forest in North China | Farmland, typical steppe |
Note: The eco-zones mentioned above are represented by codes (Zhang et al., 2013). |
Figure 1 Eco-geographical regionalization, field samples and grassland distribution in Inner Mongolia in 2010 |
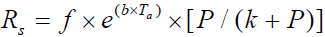
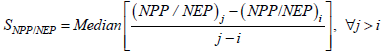
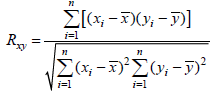
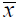 is the average NPP for years.
is the average NPP for years. 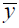 is mean precipitation or mean temperature over years (2001-2012) and n is the number of samples.
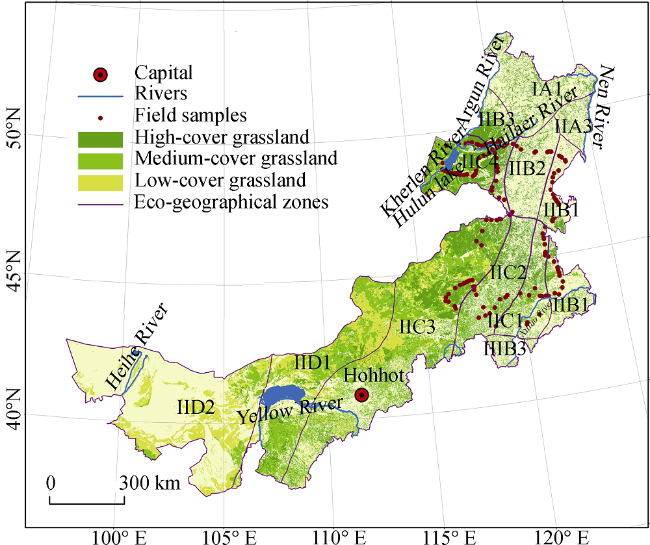
is mean precipitation or mean temperature over years (2001-2012) and n is the number of samples.Figure 2 Spatial distribution of average NPP in the Inner Mongolia grassland ecosystem from 2001 to 2012 |
Table 2 Average NPP in eco-geographical zones of the Inner Mongolia grassland ecosystem from 2001 to 2012 |
| Eco-zones | NPP (g C/m2) |
| IA1 | 590.99 |
| IIA3 | 591.27 |
| IIB1 | 405.05 |
| IIB2 | 593.64 |
| IIB3 | 487.52 |
| IIC1 | 363.16 |
| IIC2 | 466.15 |
| IIC3 | 301.88 |
| IIC4 | 323.65 |
| IID1 | 153.05 |
| IID2 | 76.45 |
| IIIB3 | 528.94 |
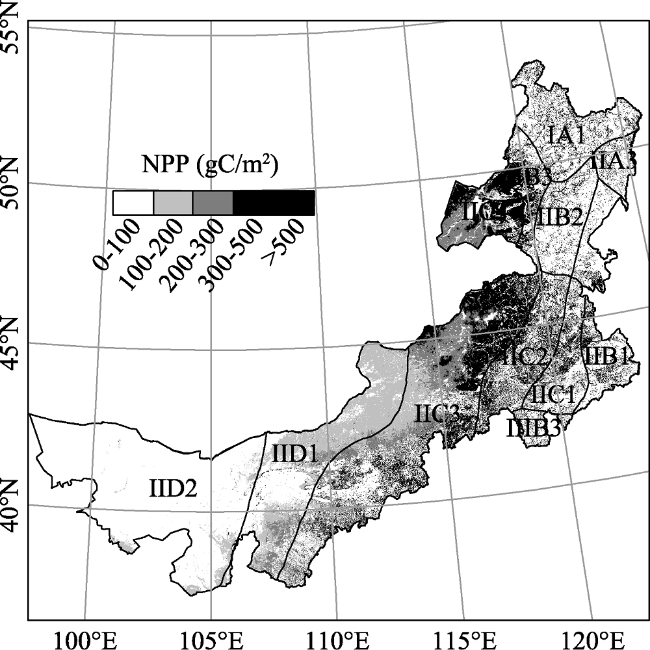
Table 3 Average NEP in eco-geographical zones of the Inner Mongolia grassland ecosystem from 2001 to 2012 |
| Eco-zones | NEP (g C/m2) |
| IA1 | 4448.90 |
| IIA3 | 3972.41 |
| IIB1 | 1297.88 |
| IIB2 | 4353.57 |
| IIB3 | 3194.29 |
| IIC1 | 885.54 |
| IIC2 | 2626.31 |
| IIC3 | 715.18 |
| IIC4 | 1295.56 |
| IID1 | -960.22 |
| IID2 | -1436.88 |
| IIIB3 | 2849.03 |
Figure 3 Spatial distribution of the total NEP in the Inner Mongolia grassland ecosystem from 2001 to 2012 |
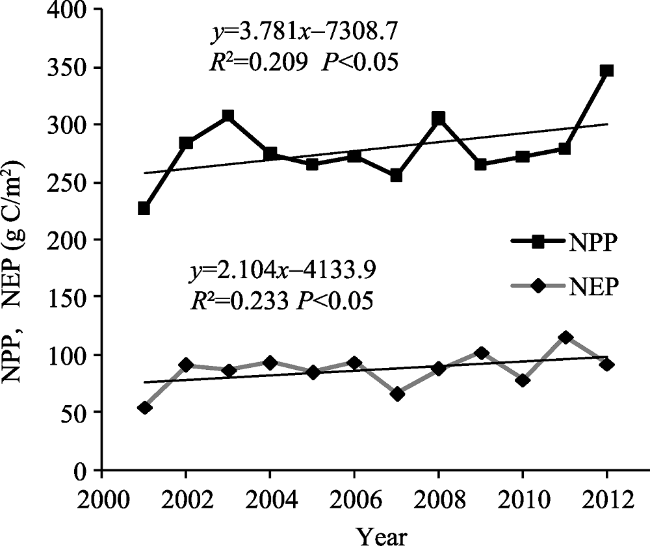
Figure 4 The variation trend in NPP and NEP in the Inner Mongolia grassland ecosystem from 2001 to 2012 |
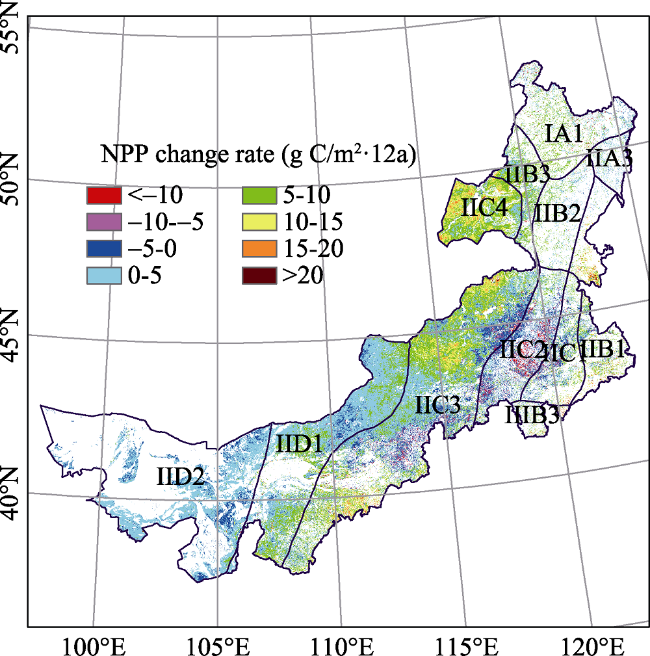
Table 4 Trend in NPP in eco-geographical zones of the Inner Mongolia grassland ecosystem from 2001 to 2012 |
| Eco-zones | SNPP |
| IA1 | 5.55 |
| IIA3 | 3.17 |
| IIB1 | 7.44 |
| IIB2 | 4.34 |
| IIB3 | 6.36 |
| IIC1 | 3.01 |
| IIC2 | -1.99 |
| IIC3 | 3.07 |
| IIC4 | 11.40 |
| IID1 | 3.04 |
| IID2 | 0.93 |
| IIIB3 | 3.73 |
Figure 5 Spatio-temporal pattern and NPP average change rate in the Inner Mongolia grassland ecosystem from 2001 to 2012 |
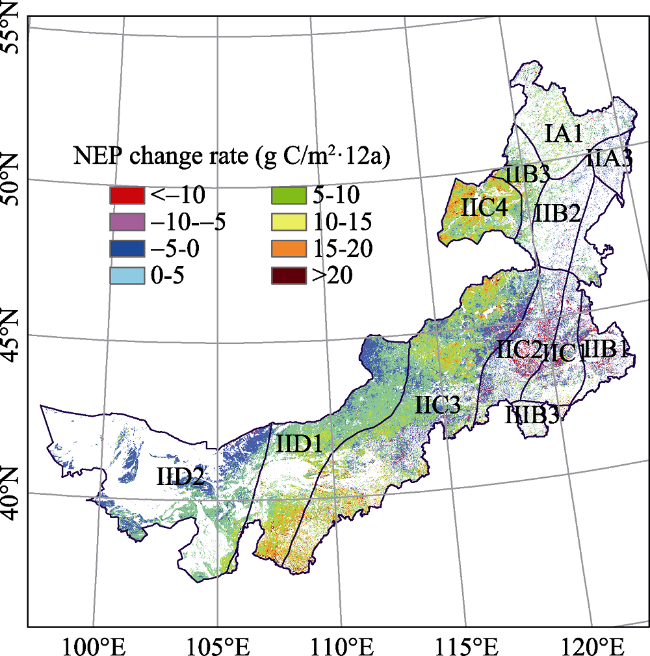
Figure 6 Spatio-temporal pattern and the NEP average change rate in the Inner Mongolia grassland ecosystem from 2001 to 2012 |
Table 5 Trend in NEP in eco-geographical zones of the Inner Mongolia grassland ecosystem from 2001 to 2012 |
| Eco-zones | SNEP |
| IA1 | 2.40 |
| IIA3 | -1.75 |
| IIB1 | -2.35 |
| IIB2 | -0.51 |
| IIB3 | 3.11 |
| IIC1 | -3.50 |
| IIC2 | -2.83 |
| IIC3 | 3.55 |
| IIC4 | 7.57 |
| IID1 | 3.35 |
| IID2 | 2.04 |
| IIIB3 | 1.06 |
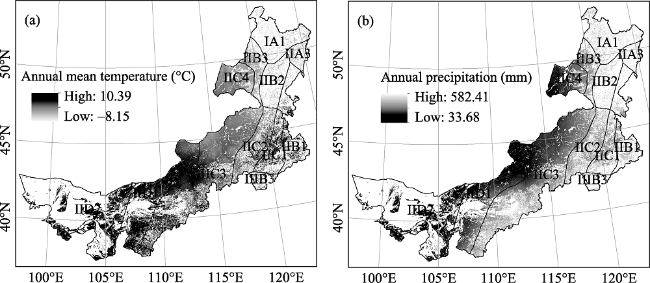
Figure 7 The spatial pattern of the annual mean temperature (a) and the annual precipitation (b) in the Inner Mongolia grassland ecosystem from 2001 to 2012 |
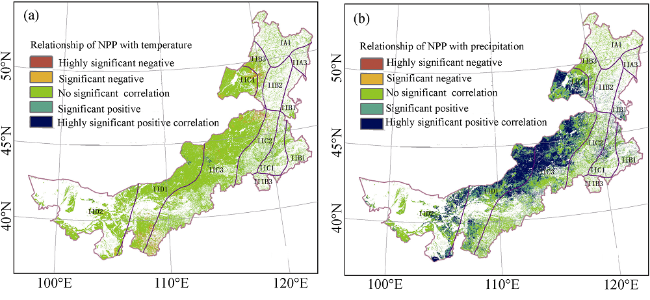
Figure 8 The significant relationship of NPP with temperature (a) and precipitation (b) in the Inner Mongolia grassland ecosystem |
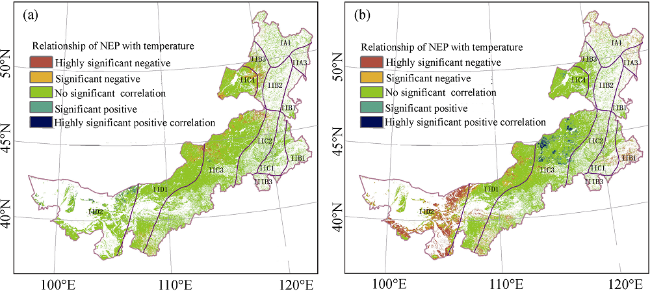
Figure 9 The significant relationship of NEP with temperature (a) and precipitation (b) in the Inner Mongolia grassland ecosystem |
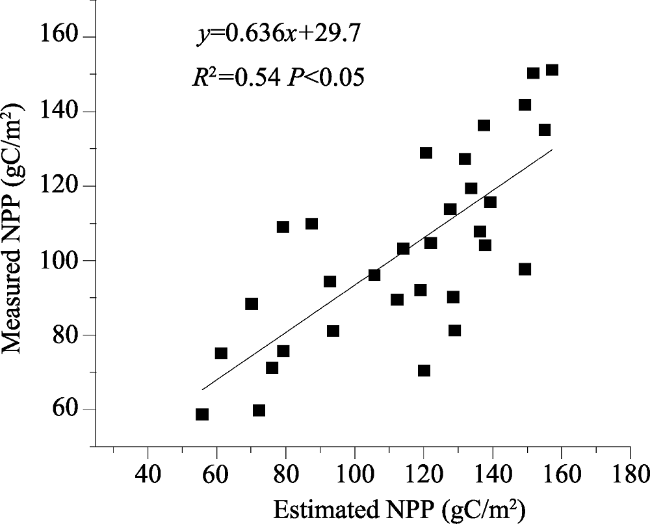
Figure 10 The relationship between the estimated and the measured values of NPP in 2011 and 2012 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}