
Journal of Geographical Sciences >
Implications of mass elevation effect for the altitudinal patterns of global ecology
Author: Zhang Baiping, Professor, specialized in physical geography and applied GIS. E-mail: zhangbp@lreis.ac.cn
*Corresponding author: Yao Yonghui, Associate Professor, E-mail: yaoyh@lreis.ac.cn
Received date: 2016-03-09
Accepted date: 2016-03-30
Online published: 2016-07-25
Supported by
National Natural Science Foundation of China, No.41421001, No.41571099, No.41030528
Copyright
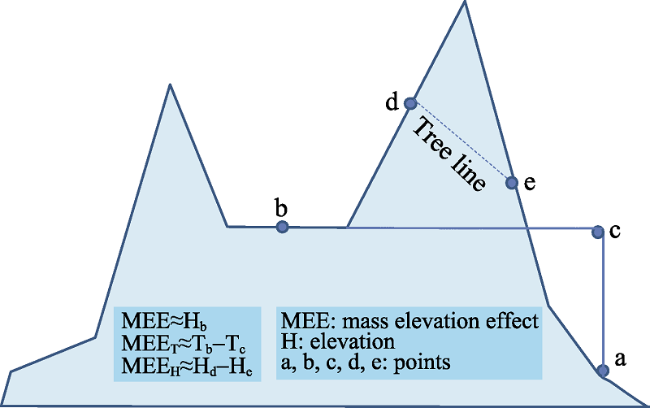
The varied altitudinal gradient of climate and vegetation is further complicated by mass elevation effect (MEE), especially in high and extensive mountain regions. However, this effect and its implications for mountain altitudinal belts have not been well studied until recently. This paper provides an overview of the research carried out in the past 5 years. MEE is virtually the heating effect of mountain massifs and can be defined as the temperature difference on a given elevation between inside and outside of a mountain mass. It can be digitally modelled with three factors of intra-mountain base elevation (MBE), latitude and hygrometric continentality; MBE usually acts as the primary factor for the magnitude of MEE and, to a great extent, could represent MEE. MEE leads to higher treelines in the interior than in the outside of mountain masses. It makes montane forests to grow at 4800-4900 m and snowlines to develop at about 6000 m in the southern Tibetan Plateau and the central Andes, and large areas of forests to live above 3500 m in a lot of high mountains of the world. The altitudinal distribution of global treelines can be modelled with high precision when taking into account MEE and the result shows that MEE contributes the most to treeline distribution pattern. Without MEE, forests could only develop upmost to about 3500 m above sea level and the world ecological pattern would be much simpler. The quantification of MEE should be further improved with higher resolution data and its global implications are to be further revealed.
ZHANG Baiping , *YAO Yonghui . Implications of mass elevation effect for the altitudinal patterns of global ecology[J]. Journal of Geographical Sciences, 2016 , 26(7) : 871 -877 . DOI: 10.1007/s11442-016-1303-2
Figure 1 A conceptual model of mass elevation effect |
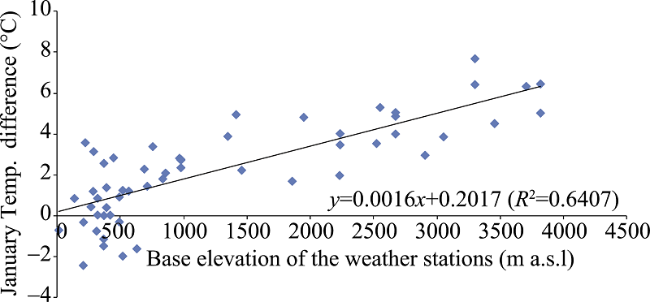
Figure 2 MEE and intra-mountain base elevation in the Andes |
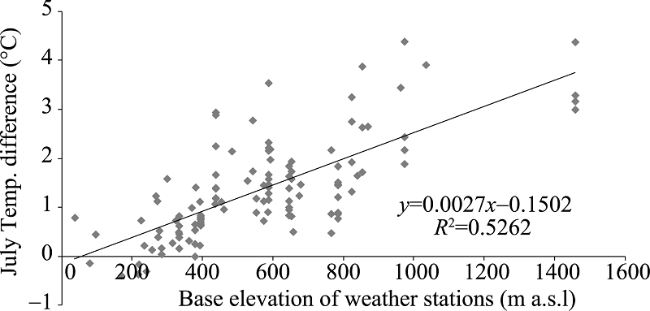
Figure 3 MEE and intra-mountain base elevation in the Alps |
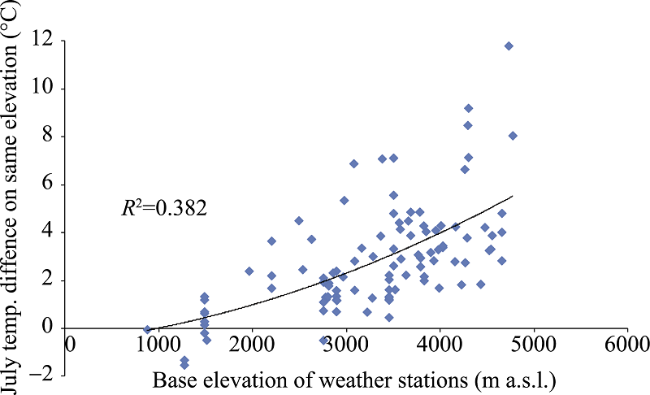
Figure 4 MEE and intra-mountain base elevation in the Tibetan Plateau |
Table 1 Contribution of MEE-influencing factors to MEE and the model coefficient of determination |
| MEE factors | Tibetan Plateau | Alps | Scandinavia | Rocky Mts. | Andes | New Zealand |
|---|---|---|---|---|---|---|
| Latitude | 38.41 | 6.19 | 17.67 | 38.52 | 4.36 | 28.30 |
| Intra-mountain base elevation | 42.66 | 56.67 | 56.10 | 61.01 | 82.80 | 32.84 |
| Hygrometric continentality | 18.93 | 37.14 | 26.23 | 0.47 | 12.84 | 38.86 |
| Model R2 | 0.515 | 0.563 | 0.476 | 0.501 | 0.646 | 0.544 |
Table 2 Distribution of the extremely high treelines in the world |
| Treeline site | Longi (°) | Lati (°) | Elev (m) | MBE (m) | Location | References |
|---|---|---|---|---|---|---|
| Nevado Sajama | -68.9 | -18.1 | 4800 | 4200 | Central Andes | Troll, 1973 |
| Baxoi county | 96.74 | 29.75 | 4900 | 4200 | S.E. Tibetan Plateau | Miehe, 2007 |
| Nyemo River (E) | 90.03 | 29.31 | 4800 | 3850 | S.E. Tibetan Plateau | Schickhoff, 2005 |
| Kyi Chu catchment | 91.60 | 30.29 | 4850 | 3800 | S.E. Tibetan Plateau | Miehe, 2007 |
| Porong Ka Monastery | 91.16 | 29.77 | 4600 | 3700 | S.E. Tibetan Plateau | Schickhoff, 2005 |
| Pamtschü | 91.96 | 29.30 | 4600 | 3650 | S.E. Tibetan Plateau | Schickhoff, 2005 |
| Nevado de Toluca | -99.7 | 19.1 | 4010 | 2800 | Southern Rocky Mts. | Körner & Paulsen, 2004 |
| Pico de Orizaba | -97.3 | 19.1 | 4020 | 2600 | Southern Rocky Mts. | Körner & Paulsen, 2004 |
| Pico de Valley | -97.3 | 19.1 | 4000 | 2600 | Southern Rocky Mts. | Hoch & Körner, 2005 |
The authors have declared that no competing interests exist.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}